Vk cc сокращение. Сервис сокращения ссылок вконтакте. Vk.cc
Согласитесь, что длинный урл адрес не всегда смотрится красиво. Как это исправить?
Уже давно существует большое количество сервисов, которые сокращают ссылки. Вы указываете оригинальный урл, и формируете сокращенный вариант. При переходе по такой ссылке, пользователь попадает на нужную страницу.
https://vk.com/ccВот так выглядит главная страница.
Скопируйте нужный урл адрес, и вставьте его в форму. Затем нажмите кнопку «Сократить» .
Вставляем ее в форму, и формируем сокращенную ссылку.
Мы получим готовый урл адрес. Он выглядит вот так:
Https://vk.cc/6huY1D
Можете скопировать его, и использовать по назначению. Попробуйте перейти по ней — вы попадете на оригинальную страницу.
Другие сервисы для сокращения ссылок
Есть большое количество аналогичных сервисов. Вы можете легко найти их через поиск.
Давайте для примера, я покажу вам такой инструмент, предоставляемый гуглом.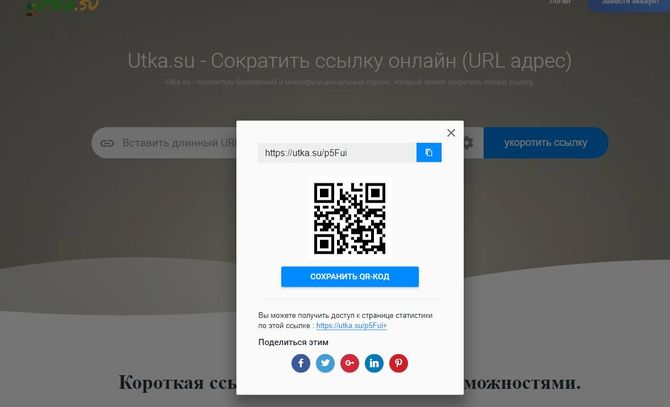
Https://goo.gl/
Откроется окно — в нем будет представлена сокращенная ссылка. Скопируйте ее и используйте.
Видео урок: как сделать сокращенную ссылку в ВК
Заключение
Если вы хотите зарабатывать на своей группе вконтакте (см. ), то вы наверняка будете использовать сокращенные ссылки. С их помощью маскируют урл партнерских программ, которые рекламируют владельцы публичных страниц.
Но это не единственный вариант. Любой длинный урл, можно переделать в короткий аналог.
Вопросы?
Вконтакте
Также он может быть полезен при попытке передать ссылку, которую социальная сеть VK.COM считает вредоносной:
Ссылка на подозрительный сайт
ВКонтакте всегда заботится о Вашей безопасности!
Часто это предупреждение не соответствует действительности и ВК просто блокирует белые сайты, для обхода этого ограничения и существуют сервисы сокращения ссылок.
Для перехода в сокращатель ссылок от вконтакте вам нужно перейти по одной из этих ссылок –
https://vk.com/cc
https://vk.сс/
На этой странице Вы можете сделать из длинной и сложной ссылки простую.
Затем нажмите “получить короткий вариант ссылки”.
Для чего можно и нужно использовать сокращение ссылок через vk.com/cc:
Не очень красивая, не правда ли? А вот как она выглядит в сокращенном виде – http://vk.cc/3HkKrK
2. Такие ссылки используются для скрытия реферальных ссылок. Чтобы не было надписи “r/18182”, “ref47474” и аналогичных реф. ссылку пропускают через сервис сокращения URL и она становится короткой и не вызывает подозрений
3. Для спама. Часто через многочисленные сервисы сокращения урлов спамеры маскируют ссылки. Сейчас вконтакте уже без проблем расшифровывает “шорт урлы” и банит тех, кто распространяет эти ссылки, даже в сокращенном виде.
5. Для увеличения места в “твите” в твиттере, так как там есть ограничение на число символов в одном посте и вставив длинную ссылку у вас может просто не остаться букв для ее описания.
Множество других способов, когда вам потребуется урезать ссылку.
На самом деле сайтов, предлагающих сократить вам ссылку очень много. Я дам вам адреса нескольких которыми пользуюсь сам:
https://bitly.com/
Если остались вопросы – спрашивайте.
Для сокращения ссылок ВК используйте официальный сервис, который находится по адресу: https://vk.com/cc или https://vk.cc и вставьте в поле нужный урл.
Рассмотрим на конкретном примере. У меня есть ссылка такого вида:
https://sayt-primer.ru/kak-sokratit-ssilku-vk.html
Если я к ней добавлю UTM – метки, то она станет еще длиннее. Метки нужны для отслеживания дополнительной статистики.
https://sayt-primer.ru/kak-sokratit-ssilku-vk.html?utm_source=Vk&utm_medium=target&utm_campaign=publik_123456&utm_content=banner_01&utm_term=allgeo
Прогоню ее через сервис и получу вот такой результат:
После сокращения ваша ссылка получит уникальный кодhttps://vk.
cc/8hr3Z3
Невозможно сократить эту ссылку (Ошибка)
Если вы видите у себя такую надпись, то значит ваш домен забанен и находится в черном списке ВК. Единственное, что вы можете сделать — это сменить домен на новый.
Если домен забанен, то появится ошибкаСтатистика
- количество уникальных посетителей и просмотров
- возраст
- географию по странам и городам
Все это будет представлено в виде удобных графиков. Тем самым, вы сможете детальней анализировать, с каких групп и пабликов по вашей ссылке лучше всего переходят.
Чтобы начать пользоваться статистикой, нажмите на иконку в виде графика как показано на скриншоте ниже.
Статистика коротких ссылок вк по разным параметрамКак посмотреть статистику конкурента (Лайфхак)
Внимательный читатель наверняка сразу же заметил галочку под полем ввода.
Если ее поставить, то статистика переходов будет видна только вам. Если проигнорировать, то доступ к статистике может получить любой желающий.
Если проигнорировать, то доступ к статистике может получить любой желающий.
Поставьте галочку, если хотите закрыть данные от конкурентов
Например, я знаю своего конкурента, который , используя продвижение через паблики, или конкурент продвигает товары из .
Все, что мне нужно про него знать — так это цифры и буквы, которые находятся в конце ссылки. Каждая ссылка имеет свой уникальный номер.
Теперь подставьте к номеру такую конструкцию:
https://vk.com/cc?act=stats&key= 8hr3Z3
После знака равно идет номер, который вас интересует. Если статистика открыта, то вы можете провести конкурентную разведку.
Статистика конкурента
На этом все!.
А знали ли Вы, что можно следить за своими конкурентами? Оставьте, пожалуйста, комментарий!
В Google считают, что сокращатель ссылок отлично справляется со своей задачей, но его время прошло. Все созданные ранее в Goo.gl ссылки продолжат работать даже после закрытия сервиса.
Однако всё равно придётся искать новый инструмент для сокращения ссылок. Вот сервисы, которые хорошо справятся с этой задачей.
1. Is.gd
Бесплатный сокращатель ссылок имеет минимум возможностей и простой интерфейс. Чтобы отслеживать статистику, не надо регистрироваться, достаточно нажать на I want to see statistics for this URL под ссылкой.
Об этом сервисе Лайфхакер уже . Отличный функциональный сокращатель с быстрым редиректом и подробной статистикой.
Сокращатель ссылок от бразильской компании Insite. Сервис порадует подробной статистикой, а также возможностью задать имя для ссылки и даже создать собственный домен. За последнее, правда, придётся заплатить.
Один из ветеранов сокращательного дела. Простой и удобный сервис, который служит верой и правдой уже много лет.
5. Ow.ly
Сервис от платформы отложенного постинга Hootsuite. Чтобы воспользоваться сокращателем, придётся зарегистрироваться в сервисе. Если вы планируете пользоваться им только для сокращения ссылок, выбирайте бесплатный тариф.
Если вы планируете пользоваться им только для сокращения ссылок, выбирайте бесплатный тариф.
6. U.to
Простой русскоязычный сервис. Чтобы посмотреть статистику, придётся зарегистрироваться. Можно обойтись и без этого, если просто хотите сократить ссылку.
Этот сервис примечателен тем, что в нём можно создавать сразу несколько ссылок.
8. Bitly
Популярный сервис с удобным интерфейсом и функциональной панелью администрирования, позволяющей отлеживать клики по ссылкам и просматривать расширенную статистику по ним.
9. Zapier URL Shortener
Отличный сервис одноименной компании для интеграции веб-приложений. Умеет не только сокращать ссылки, но и делать это автоматически при срабатывании определенных триггеров.
10. Rebrandly
Продвинутый сервис, который пригодится для продвижения бренда. С помощью Rebrandly можно создавать персонализированные ссылки с доменом вашего сайта.
11.
 Lnnk
LnnkОчень простой сервис сокращения, в котором можно генерировать защищенные паролем ссылки. Lnnk работает как в качестве веб-сервиса, так и в виде браузерного расширения.
12. Polr
Сервис Polr позволит быстро сократить любую ссылку и настроить ее отображение, выбрав окончание на свое усмотрение. Кроме того можно воспользоваться исходным кодом Polr и сделать собственную сокращалку.
Как делать ссылки vk cc : Радиосхема.ру
Которые помогут заменить Google URL Shortener после его закрытия.
13 апреля 2018 года Google начнёт ограничивать доступ к сервису goo.gl, а 30 марта 2019 года окончательно прекратит поддержку платформы. Рассказываем о восьми сервисах для сокращения ссылок, которые могут стать альтернативой Google URL Shortener.
«Кликер» преобразует ссылки в формат clck.ru и создает QR-код, который можно сохранить как изображение. Сервис также дублирует короткую ссылку, прописывая её по буквам.
Чтобы воспользоваться vk. cc, необходимо авторизоваться через «ВКонтакте». Сервис позволяет сокращать ссылки, а также отслеживать статистику по ним: число переходов, уникальных посетителей и просмотров.
cc, необходимо авторизоваться через «ВКонтакте». Сервис позволяет сокращать ссылки, а также отслеживать статистику по ним: число переходов, уникальных посетителей и просмотров.
Cокращать ссылки на Bitlу можно анонимно, но чтобы получить доступ к инструментам аналитики, придётся зарегистрироваться. Авторизованные пользователи также могут изменять ссылки — например, создавать собственные короткие имена.
Ow.ly принадлежит и расположена на платформе управления публикациями в соцсетях Hootsuite. Сервисом можно воспользоваться, зарегистрировавшись через электронную почту или аккаунт в соцсетях. Авторизованные пользователи получают бесплатный пакет на 30 дней на Hootsuite.
Сервис to.click позволяет сокращать и редактировать ссылки и анализировать поступающий по ним трафик. Ссылками можно управлять пр помощи API, кроме того, to.click предлагает сокращать ссылки при помощи бота в Telegram.
Сервис для сокращения ссылок от конструктора сайтов uCoz. Для того, чтобы получить доступ к статистике переходов и функции редактирования ссылок, придётся зарегистрироваться.
Droplr – это облачный сервис для обмена фото, видеофайлами и другими файлами. Платформа также позволяет сокращать ссылки — для этого нужно авторизоваться на сайте при помощи электронной почты.
Ну как, зачем? Goo.gl такой удобный. Много есть не просит зачем его убивать( не понимаю(((
urlog.ru юзайте, сейчас самая топовая сокращалка! Вот такие визитки с QR-Code генерирует автоматически, для соцсетей и других известных сайтов логотип меняется тоже, думаю для людей занимающихся бизнесом будет полезно!
Удобный-то , он да, но в вк не проходит. А Глобус, укоротить не получается в вк. Что делать в такой ситуации, может кто-нить встречался с такой проблемой и решил?
«Кликер» от «Яндекса»
Оказывается при загрузке видео урезается звук и конвертируется в гиф 🙁
Сюрпризы, сюрпризы . новые фичи добавляются и исчезают, и блога разработчиков vc.ru нету, чтобы о них прочитать 🙁
Почему Яндекс не может сделать просто сервис . Почему он вечно такой не удобный и перегруженый.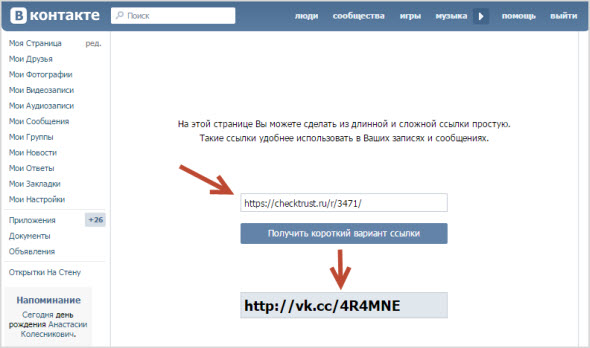 Так ещё с интерфейсом как 20-летней давности.
Так ещё с интерфейсом как 20-летней давности.
Гуглы — козлы. Нахрена закрывать сервис?
Активно распространяют вирусы и фишинговые страницы. Модерировать им лень. Зарабатывать на этом сложно — хотя могли бы при редиректе добавлять страницу с адресом куда человек собрался, предупреждением о небезопасности перехода, и показывать рекламу. Еще могли бы всю базу ссылок шерстить антивирусом нон-стоп — он у них есть, в почте работает ведь.
Но им проще закрыть.
А еще есть https://ylink.pro. С ботом в Telegram (https://t.me/ylinkpro_bot), умеющим работать в инлайн режиме, чатах, каналах и хранить картинки
Из представленный самый полезный u.to, так как имеет меньше всего символов. Это важно, что сокращении текста сообщений, например, в телеграме (ограничения в 200 символах при медиа)
Приятно, что наш сервис https://to.click попал в этот обзор. Могу ответить на вопросы и помочь с переездом если нужно.
Sergey Boyarsky, jr.
Две недели пользования вашим сервисом для переадресации на сайт регистрации на некоммерческий корпоративный ивент в google sites.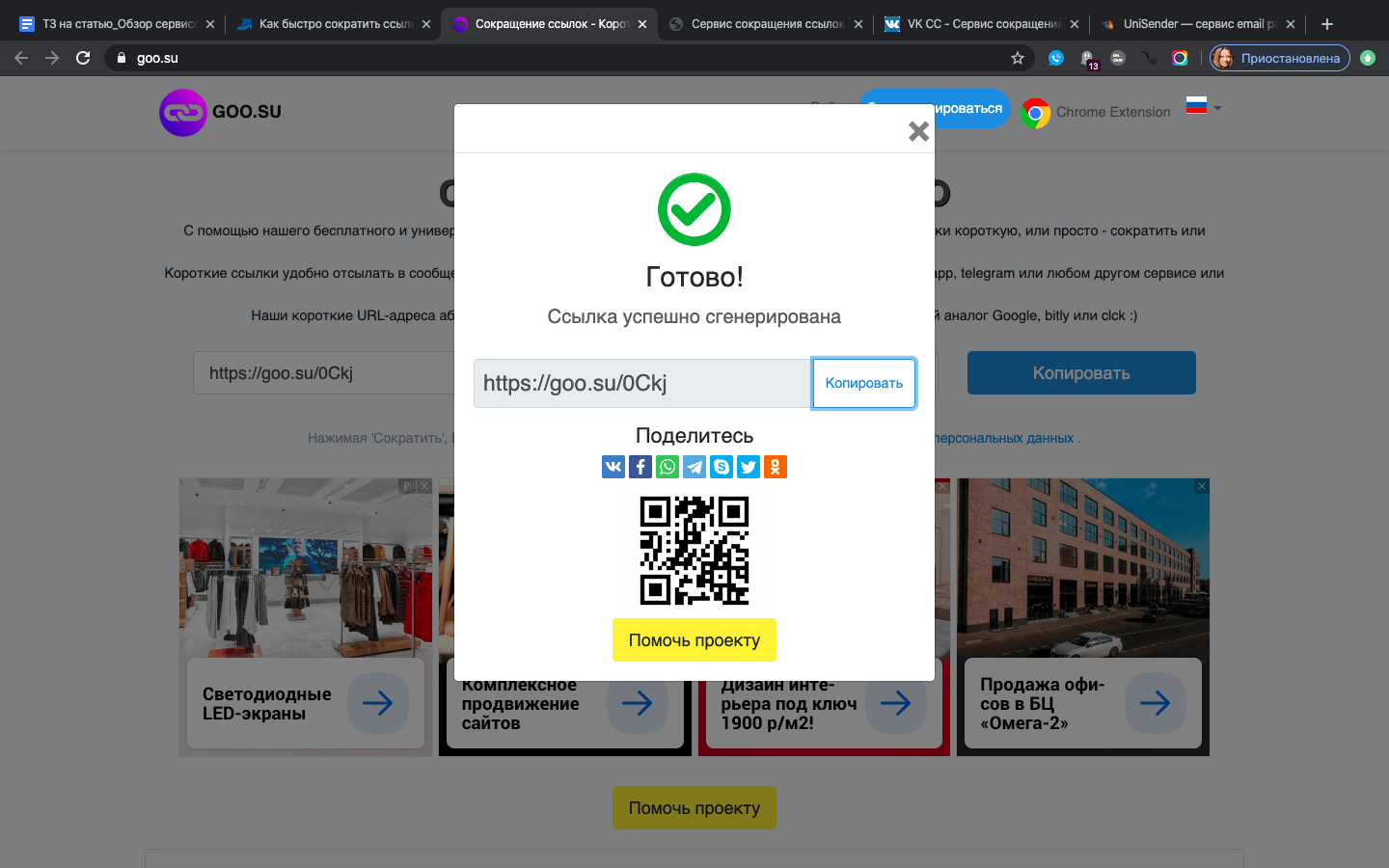 Как результат: каждый день сервис по три-пять раз выпадает в ошибку и недоступен каждый раз по 10-20 минут. И как апогей — моя ссылка без предупреждения заблокирована, хотя она не нарушает ни одно из правил. Достучаться в поддержку просто нереально. Такой сервис отправляется в топку.
Как результат: каждый день сервис по три-пять раз выпадает в ошибку и недоступен каждый раз по 10-20 минут. И как апогей — моя ссылка без предупреждения заблокирована, хотя она не нарушает ни одно из правил. Достучаться в поддержку просто нереально. Такой сервис отправляется в топку.
если нужно сокрашать ссылки для апп сторов и умные редиректы делать то еще есть https://onsto.re/
куда лучше использовать Российский продукт https://utka.su/
Еще хороший сервис сокращения ссылок https://ogo.gl/
Есть статистика, QR-коды, можно поставить лимит на кол-во переходов ,а также есть возможность самому выбрать текст сокращенной ссылки
https://vk-cc.ru пользуюсь для сокращения со своим доменом или https://vk.io, по функционалу как битли, только русское и бесплатное. Тоже можно свой домен подключить и тоже можно хитрые редиректы делать
Thanks for a mention about Droplr guys!
We are proud to be on your list! I think it’s good to say in Droplr you can customize your links (if you have a PRO account) and send the link from your own domain like yourdoma. in/1234 you can send share files, photos, texts or even links 🙂
in/1234 you can send share files, photos, texts or even links 🙂
Best,
Filip from Droplr
Неплохие сервисы в списке, пользовался раньше кликером, но сейчас мне понравился другой сервис по сокращению ссылок, которым я пользуюсь уже на протяжении месяца urlog.ru, странно что его нет в списке,я нашел его у другого блогера в списке, не зря он стоял у него на первом месте. Очень хороший сервис, есть все инструменты для сокращения ссылок, будь то пароль на ссылку или графики переходов, чего стоит одна стоуровневая проверка, это значит что ваша короткая ссылка ни когда не совпадет с чей либо и самое главное что urlog бесплатный. Например ссылку на эту статью: https://vc.ru/flood/35599-7-servisov-dlya-sokrashcheniya-ssylok он сократит вот так: https://urlog.ru/u84ep3 легко запоминается и ничего лишнего! Еще понравилось копирование сгенерированный ссылки в один клик и оптимизированый под мобильные устройства интерфейс, очень выручает когда медленный интернет загружается сайт на ура, в общем рекомендую!
0
Неплохие сервисы в списке, пользовался раньше кликером, но сейчас мне понравился другой сервис по сокращению ссылок, которым я пользуюсь уже на протяжении месяца urlog. ru, странно что его нет в списке,я нашел его у другого блогера в списке, не зря он стоял у него на первом месте. Очень хороший сервис, есть все инструменты для сокращения ссылок, будь то пароль на ссылку или графики переходов, чего стоит одна стоуровневая проверка, это значит что ваша короткая ссылка ни когда не совпадет с чей либо и самое главное что urlog бесплатный. Например ссылку на эту статью: https://vc.ru/flood/35599-7-servisov-dlya-sokrashcheniya-ssylok он сократит вот так: https://urlog.ru/u84ep3 легко запоминается и ничего лишнего! Еще понравилось копирование сгенерированный ссылки в один клик и оптимизированый под мобильные устройства интерфейс, очень выручает когда медленный интернет загружается сайт на ура, в общем рекомендую! Что то тормозят комментарии!
ru, странно что его нет в списке,я нашел его у другого блогера в списке, не зря он стоял у него на первом месте. Очень хороший сервис, есть все инструменты для сокращения ссылок, будь то пароль на ссылку или графики переходов, чего стоит одна стоуровневая проверка, это значит что ваша короткая ссылка ни когда не совпадет с чей либо и самое главное что urlog бесплатный. Например ссылку на эту статью: https://vc.ru/flood/35599-7-servisov-dlya-sokrashcheniya-ssylok он сократит вот так: https://urlog.ru/u84ep3 легко запоминается и ничего лишнего! Еще понравилось копирование сгенерированный ссылки в один клик и оптимизированый под мобильные устройства интерфейс, очень выручает когда медленный интернет загружается сайт на ура, в общем рекомендую! Что то тормозят комментарии!
Wlinks.ru короткие ссылки с паролем
https://ur-l.ru — очень хороший и удобный бесплатный сервис сокращения ссылок.
Есть статистика переходов, произвольное название ссылки, QR-код, защита паролем, срок действия ссылки, геотаргетинг, панель управления и многое другое.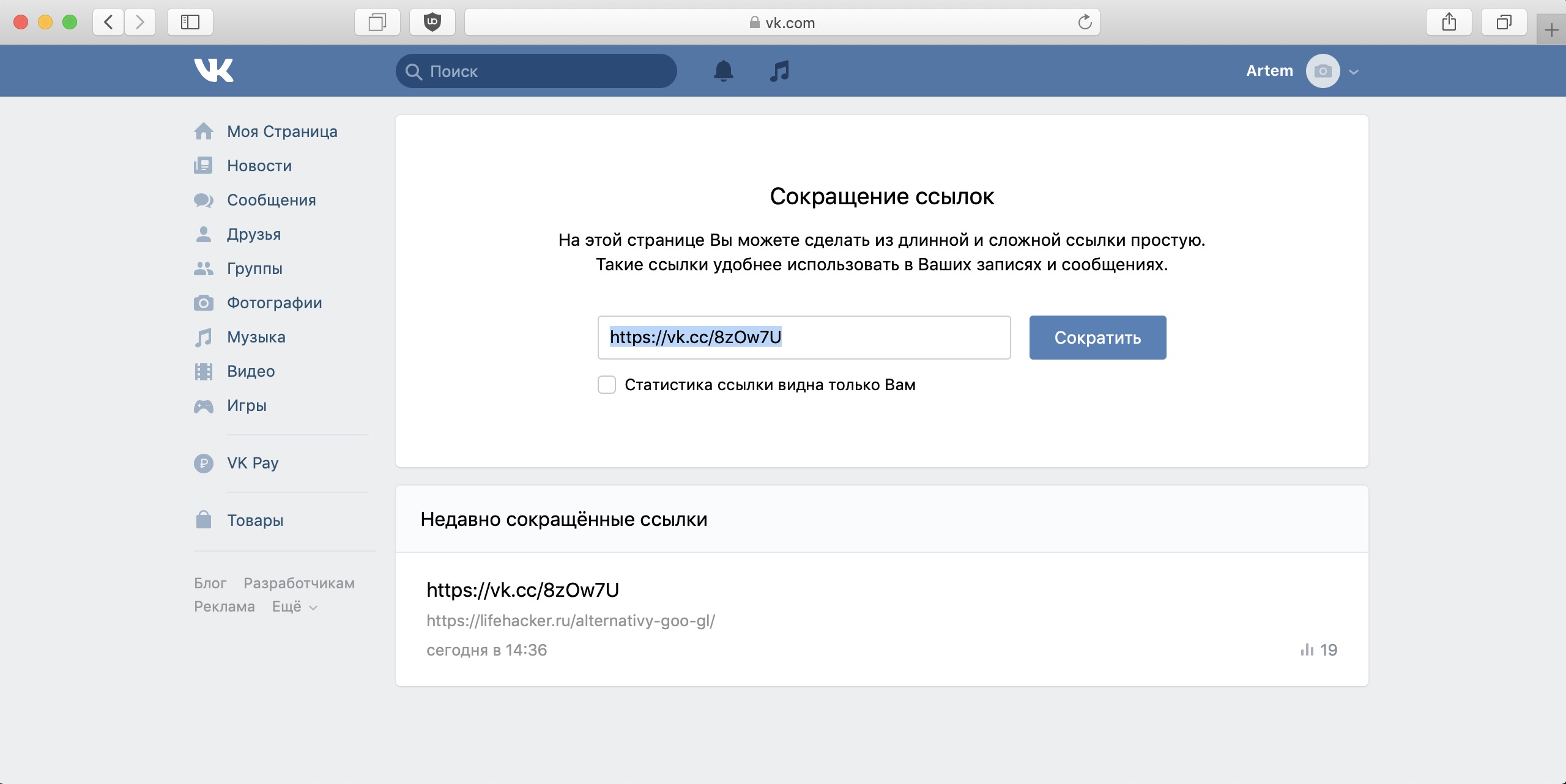
А чего в списке нет https://urlog.ru вполне достойный сервис, всяким bitly и vk Мне нравится как он делает визитки с QR-code, вот например короткая ссылка на эту статью https://urlog.ru/u84ep3 и визитка на неё.
битлу всем хорош, но довольно часто он почему-то баниться Роскомнадзором.
Отличные новости для тех, кто работает с публикациями, занимается рекламными метриками или просто не любит громоздкие ссылки.
Мы прокачали vk.cc — наш сервис для сокращения ссылок. Теперь он умеет считать статистику переходов и отдавать информацию через API.
Зачем нужен vk.cc?
Чтобы превращать длинные и неуклюжие ссылки в короткие и аккуратные.
До:
www.cybersport.ru/news/skripty-dlya-botov-dobavleny-v-workshop-dota-2
После:
vk.cc/6iXcdC
Такие ссылки радуют глаз и отлично вписываются в любой текст:
Новый vk.cc помнит всё, что Вы сокращали. В списке быстро найдётся нужный адрес:
Справа — число переходов.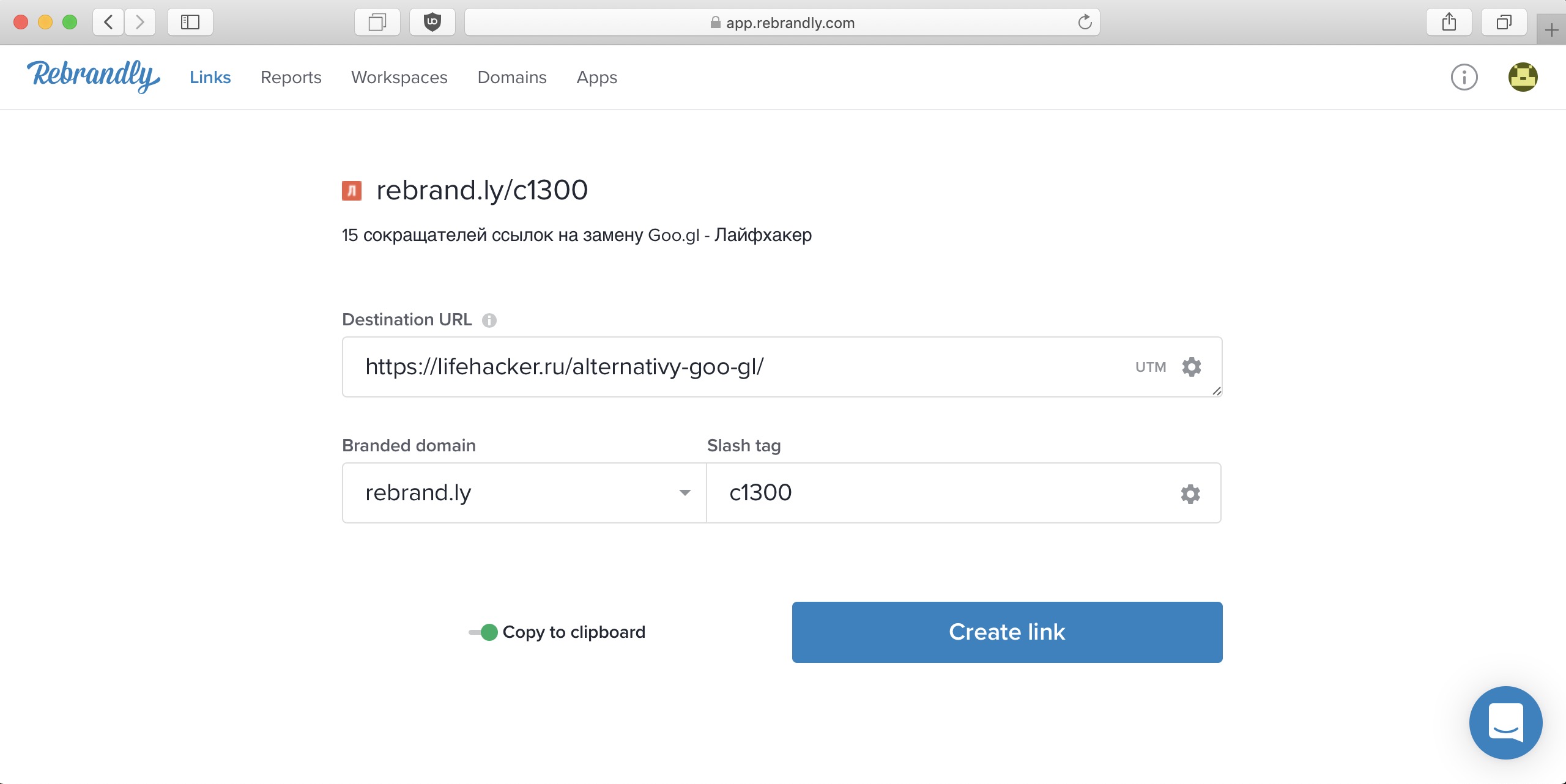 Нажмите на него, чтобы открыть статистику.
Нажмите на него, чтобы открыть статистику.
Статистика
Изучайте подробную статистику переходов: платформы, география, пол и возраст посетителей. Где бы ни оказалась Ваша ссылка, мы посчитаем честные данные по визитам пользователей ВКонтакте. Да, даже если она опубликована за пределами vk.com.
Значения из красивых графиков и диаграмм можно выгрузить в таблицу Excel или .csv-файл. Для особых ценителей мы добавили немного API.
Автоматизируйте сокращение ссылок и получайте статистику напрямую из своего приложения — теперь это доступно с помощью специальных методов.
Подробнее о них в нашей документации:
Возможно вы уже сталкивались с проблемой, когда нужно отправить другу ссылку, а вконтакте блокирует ее. Или другая ситуация — вы хотите разместить ссылку в публичном доступе, а она длинная как простыня и выглядит некрасиво. В обоих случаях на помощь приходит сокращение ссылок ВК.
В этой статье мы рассмотрим 3 популярных сервиса для сокращения ссылок: vk. cc, Кликер и u.to, их плюсы и минусы. Попробуем сократить в них разные ссылки и выберем подходящий.
cc, Кликер и u.to, их плюсы и минусы. Попробуем сократить в них разные ссылки и выберем подходящий.
Содержание
Сервис коротких ссылок vk.cc
Сокращатель ссылок vk.cc доступен по 2 ссылкам vk.cc и vk.com/cc. Обе ссылки ведут на одну и ту же страницу.
В пустое поле нужно вставить ссылку, которую вы хотите сократить и нажать кнопку «Сократить».
После сокращения готовая короткая ссылка появится в этом же поле. Вам останется только скопировать ее и разместить там, где хотели или отправить в сообщении другу.
Через сервис коротких ссылок vk.cc можно просматривать статистику по сгенерированным ссылкам. В статистике можно увидеть количество переходов по вашей ссылке.
Чтобы посмотреть статистику нужно щелкнуть в область, отмеченную на скриншоте ниже. Если щелкнуть на саму ссылку, то произойдет переход по ней, а не в статистику.
Можно посмотреть количество переходов по ссылке и не заходя в статистику, она видна в нижем левом углу строки.
Просмотр статистики
Просмотр статистики может понадобиться, если вы разместили вашу реферальную ссылку в какой-нибудь группе, например, и хотите посмотреть сколько людей по ней перешло, чтобы оценить, насколько эффективно было ваше объявление.
Важно! Если не поставить галочку «Статистика видна только Вам», то ее смогут посмотреть все желающие и тоже оценить эффективность вашей рекламы. Так что не забывайте ставить, если не хотите делиться с конкурентами информацией.
Все ранее сокращенные вами ссылки будут доступны на странице vk.cc в разделе «Недавно сокращенные ссылки», если вы будете залогинены в вашем аккаунте.
Одно но, этот сервис не будет сокращать ссылки, которые ему не нравятся.
А не нравится ему все, что связано со спамом, со сторонними сервисами накрутки, сомнительным заработком. Т.е. скорее всего реферальные ссылки сократить не получится.
Если вы получили сообщение «Невозможно сократить эту ссылку», то это значит, что ваша ссылка как раз попадает в список неугодных ВК.
Плюсы и минусы сервиса vk.cc
- Можно перейти прямо из Вконтакта
- Доступен просмотр статистики
- Не все ссылки можно сократить
- Требуется регистрация Вконтакте
Вывод: Сервис хорош для сокращения длинных ссылок, например, с utm-метками, подойдет для таргетологов и смм-менеджеров. Сокращает не все ссылки.
Как сократить ссылки через Кликер
Еще один сокращатель ссылок принадлежит Яндексу и хорош тем, что не придирается к ссылкам и сокращает любые ссылки без разбора.
Находится по адресу clck.ru.
Интерфейс предельно прост: вставляете свою ссылку и нажимаете клик.
Статистики этот сервис не дает, но зато генерирует QR-код.
Для тех, кто не знает, поясню, что QR-код можно отсканировать через сканер мобильного телефона и получить из него ссылку, чтобы не набивать ее руками в телефоне.
Плюсы и минусы сокращателя ссылок Кликер
- Простой интерфейс
- Не требует регистрации
- Генерирует QR-код
- Сокращает все ссылки без ограничений
- Не отдает и не хранит статистику переходов
Сокращение ссылок через u.
 to
toЕще один простой сервис сокращения ссылок.
Сервис хорош тем, что хранит и отдает статистику переходов по ссылке. Есть возможность зарегистрироваться и просматривать данные через личный кабинет.
Если хотите просматривать статистику, то лучше зарегистрироваться, т.к. сервис хоть и помнит ваши ссылки без регистрации, но после очистки куки все забудет.
Регистрация в u.to
Для регистрации нужно в верхнем правом углу экрана нажать ссылку Регистрация. Откроется форма ввода данных, которую нужно заполнить полностью!
Рекомендую в форме регистрации под Именем, Фамилией и Датой рождения в строках Могут видеть: Все заменить на никто. Ну незачем всем видеть персональные данные, как мне кажется.
После заполнения нажимаете на кнопку «Создать аккаунт», заходите на электронную почту, которую указали и переходите по ссылке для подтверждения аккаунта.
Все, теперь можно заходить в личный кабинет.
Через личный кабинет доступна полная статистика ссылок, возможность создания сокращенных, таргетированных ссылок и дип линков.
И самая интересная возможность, это делать сокращенные ссылки с привязкой к своем домену, если он у вас есть.
Так что пожалуй этот сервис можно признать одним из лучших и по простоте и по функционалу.
Плюсы и минусы сервиса u.to:
- Не требует обязательной регистрации;
- Можно создать личный кабинет;
- Сокращает все ссылки;
- Делает таргетированные ссылки и дип линки.
Часто при добавлении ссылок в посты ВКонтакте они получаются очень длинными и могут занимать значительную часть контента. Длинная ссылка смотрится некрасиво и многих пользователей это раздражает. Для сокращения ссылок ВКонтакте создали специальный сервис, который открывается по следующим ссылкам: vk.cc и vk.com/cc . Достаточно вставить вашу длинную ссылку в поле для сокращения, выбрать кому видна статистика (только вам или всем) и нажать кнопку сократить.
Сократитель ссылок vk.cc
В итоге вы получаете короткую ссылку. При переходе по этой ссылке пользователь будет перенаправлен на вашу ссылку, которую вы сокращали в vk.cc . По каждой сокращенной ссылке доступен просмотр статистики. Статистика показывает число переходов по датам и распределение аудитории по полу и возрасту. Теперь поделимся с вами лайфхаком как посмотреть статистику по ссылкам конкурентов. Если хозяин не закрыл доступ к статистике, то любой пользователь сможет посмотреть статистику по следующей ссылке: vk.com/cc?act=stats&key=aaKncr , где key = символы, которые следуют после vk.cc/ ссылки вашего конкурента.
При переходе по этой ссылке пользователь будет перенаправлен на вашу ссылку, которую вы сокращали в vk.cc . По каждой сокращенной ссылке доступен просмотр статистики. Статистика показывает число переходов по датам и распределение аудитории по полу и возрасту. Теперь поделимся с вами лайфхаком как посмотреть статистику по ссылкам конкурентов. Если хозяин не закрыл доступ к статистике, то любой пользователь сможет посмотреть статистику по следующей ссылке: vk.com/cc?act=stats&key=aaKncr , где key = символы, которые следуют после vk.cc/ ссылки вашего конкурента.
Также ВКонтакте добавили автоматическую генерацию QR-кода сокращенной ссылки. QR-код можно скачать в формате PNG или SVG.
QR-код сокращенной ссылки в vk.cc
Советуем обязательно пользоваться сократителем ссылок для ВК при создании рекламных кампаний. Так рекламные посты выглядят аккуратно и читабельно. Ну а если вам понадобится сочная горячая целевая аудитория наш парсер ВКонтакте к вашим услугам 😉
Сокращение ссылок в Гугл (goo.
 gl) и ВК (vk.cc) — какой сократитель …
gl) и ВК (vk.cc) — какой сократитель …Здравствуйте, уважаемые читатели блога KtoNaNovenkogo.ru. Давайте сразу попробуем ответить на вопрос — зачем может понадобиться сокращение (укорачивание) ссылок? На самом деле вариантов много.
Например, короткие ссылки намного удобнее будет отправить через SMS или сервис мгновенных сообщений, чем исходный Урл, который может содержать сотни символов (набирать замучаетесь). Правда в этом случае может помочь преобразование Урлов в QR-коды, которые будет легко считать с мобильного телефона, но лучше уж подстраховаться.
Кроме этого некоторые сервисы (типа Твиттера) не допускают написания длинных сообщений, поэтому сокращение длины вставляемой ссылки может быть очень важным фактором. Да, собственно, Твиттер и стал отправной точкой для набора популярности разных онлайн-сократителей. Правда Twitter процесс укорачивания уже давно автоматизировал (длинный Урл преобразуется в короткий без вашего участия).
Но не только это. В большинстве сервисов сокращенных ссылок ведется подробная и детальная статистика переходов, что может быть полезно для отслеживания тех или иных процессов. Ну и, конечно же, партнерские программы с их реферальными ссылками. Кто не знает, что это такое — читайте в приведенной статье, а кто уже давно зарабатывает на партнерках, понимает как важно уберечь окончание рефссылки в целости и сохранности, ибо именно оно позволяет нам зарабатывать.
В большинстве сервисов сокращенных ссылок ведется подробная и детальная статистика переходов, что может быть полезно для отслеживания тех или иных процессов. Ну и, конечно же, партнерские программы с их реферальными ссылками. Кто не знает, что это такое — читайте в приведенной статье, а кто уже давно зарабатывает на партнерках, понимает как важно уберечь окончание рефссылки в целости и сохранности, ибо именно оно позволяет нам зарабатывать.
Сокращатель ссылок от Гугла (goo.gl) и его возможности
Официально этот сервис коротких ссылок называется Google URL Shortener, но его больше узнают по его довольно таки забавному Урл адресу, а точнее доменному имени (что это такое?) — goo.gl.
Самым главным его достоинством является то, что он принадлежит Гуглу, в результате чего можно констатировать и его основные преимущества перед конкурентами по укорачиванию Урлов:
- Вы должны понимать, что пользователь, кликая по любой короткой ссылке, попадает на нужную страницу не сразу, а транзитом через сайт (веб-сервер) сервиса-сократителя.
 А если таких переходов будет совершаться огромное количество одновременно? Правильно, сервер может начать подтормаживать. Будут ли люди, переходящие по вашим укороченым ссылкам, ждать долго? Не факт.Поэтому очень важна надежность и нагрузочная способность серверов подобных сервисов. А кто по вашему обладает самой мощной серверной мощностью в мире? Правильно, Гугл. И поэтому именно goo.gl, при прочих равных условиях, будет предпочтительнее выбирать из-за высокой гарантииработоспособности сокращенных в Google URL Shortener ссылок.
А если таких переходов будет совершаться огромное количество одновременно? Правильно, сервер может начать подтормаживать. Будут ли люди, переходящие по вашим укороченым ссылкам, ждать долго? Не факт.Поэтому очень важна надежность и нагрузочная способность серверов подобных сервисов. А кто по вашему обладает самой мощной серверной мощностью в мире? Правильно, Гугл. И поэтому именно goo.gl, при прочих равных условиях, будет предпочтительнее выбирать из-за высокой гарантииработоспособности сокращенных в Google URL Shortener ссылок. - Для получения короткой ссылки в goo.gl вам не нужно будет регистрироваться, ибо будут использоваться ваши данные из уже существующего Google аккаунта, который имеется у всех, кто пользуется почтой Джимайл (с высокой степенью защищенности), зарабатывает в Гугл Адсенсе, установил код счетчика Аналитикса, использует сервис поиска упоминаний Google Alerts, покупает рекламу в Адвордсе и т.п.
- Также очень важной проблемой при работе с сокращателями является возможность прекращения работы коротких ссылок через какое-то время. Согласитесь, что постоянно мониторить их на работоспособность не комильфо. Гугловский же сократитель goo.gl хранит короткие ссылки в целости до тех пор, пока существует ваш аккаунт. Конечно же, есть риск, что данный сервис будет закрыт (такое иногда случается, например, с Гугл ридером), но вы по крайней мере будете заранее об этом предупреждены.
 А если таких переходов будет совершаться огромное количество одновременно? Правильно, сервер может начать подтормаживать. Будут ли люди, переходящие по вашим укороченым ссылкам, ждать долго? Не факт.Поэтому очень важна надежность и нагрузочная способность серверов подобных сервисов. А кто по вашему обладает самой мощной серверной мощностью в мире? Правильно, Гугл. И поэтому именно goo.gl, при прочих равных условиях, будет предпочтительнее выбирать из-за высокой гарантииработоспособности сокращенных в Google URL Shortener ссылок.
А если таких переходов будет совершаться огромное количество одновременно? Правильно, сервер может начать подтормаживать. Будут ли люди, переходящие по вашим укороченым ссылкам, ждать долго? Не факт.Поэтому очень важна надежность и нагрузочная способность серверов подобных сервисов. А кто по вашему обладает самой мощной серверной мощностью в мире? Правильно, Гугл. И поэтому именно goo.gl, при прочих равных условиях, будет предпочтительнее выбирать из-за высокой гарантииработоспособности сокращенных в Google URL Shortener ссылок. Согласитесь, что постоянно мониторить их на работоспособность не комильфо. Гугловский же сократитель goo.gl хранит короткие ссылки в целости до тех пор, пока существует ваш аккаунт. Конечно же, есть риск, что данный сервис будет закрыт (такое иногда случается, например, с Гугл ридером), но вы по крайней мере будете заранее об этом предупреждены.
Согласитесь, что постоянно мониторить их на работоспособность не комильфо. Гугловский же сократитель goo.gl хранит короткие ссылки в целости до тех пор, пока существует ваш аккаунт. Конечно же, есть риск, что данный сервис будет закрыт (такое иногда случается, например, с Гугл ридером), но вы по крайней мере будете заранее об этом предупреждены.Работать с укорачивателем Гоо.гл до смешного просто. Вводите в приведенную форму Урл (универсальный идентификатор ресурса), который требуется сократить, после чего жмете на кнопку «Shorten URL» и тут же получаете возможность скопировать получившуюся ссылку, начинающуюся с доменного имени goo.gl:
В открывшемся окне вы увидите скриншот страницы, адрес которой укорачивается, а чуть выше будет приведена и сама короткая ссылка. Чтобы ее скопировать в буфер обмена достаточно будет нажать на пиктограммку листочков бумаги, расположенную справа от нее. После этого всплывающее окно можно будет закрыть и увидеть список всех ссылок, которые вы когда-либо сокращали в сервисе Гоо. гл под данным своим аккаунтом:
гл под данным своим аккаунтом:
На этой же странице можно будет посмотреть и число переходов по сокращенным в этом сервисе ссылкам, а при подведении курсора мыши к строчке с нужным «укоротом», можно будет при необходимости повторно скопировать этот короткий Урл в буфер обмена. Также справа появится выпадающее меню, где кликнув по пункту «Analytics Data» можно будет статистику кликов изучить во всех подробностях:
Там же вы сможете скопировать картинку с QR-кодом для того, чтобы данную ссылку можно было бы быстро перенести на мобильный телефон (с помощью программ распознавания такого вида кодов — фотографируете его с телефона и тут же попадаете на страницу сайта, куда это Урл ведет). Ну, и само собой, что ставшие ненужными короткие ссылки можно будет из этого списка скрыть с помощью пункта «Hide URL» выпадающего меню.
Из недостатков сервиса-сократителя Goo.Gl можно отметить:
- Все укороты собраны в одну кучу и нет никакой возможности распихать их по папкам.
- Тут не получится сделать несколько разных коротких ссылок ведущих на одну и ту же страницу, например для тестирования их расположения и выбора наиболее эффективных мест на сайте.
- Так же осторожно стоит использовать короткие ссылки из Гоо.гл в почтовых рассылках, ибо эти укороты могут вдруг перестать работать из-за подозрений в распространении спама.

Где еще можно надежно укоротить ссылку?
- Vk.cc (vk.com/cc) — официальный сокращатель ссылок от ВК. Однако, созданные в нем короткие Урлы можно использовать не только внутри этой социальной сети, но и в любом другом месте. Работает он предельно просто: вставляете Урл, который требуется сократить, жмете на кнопку и копируете короткий вариант ссылки. И все! Правда никакой статистики и даже списка уже созданных ранее вами сокращенных Урлов нет (придется самим у себя на компе создавать и хранить их базу, что не очень удобно).
Зато не возникает вопросов с работоспособностью укороченных в ВК ссылок, ибо его серверные мощности просто огромны.
Да и регистрации никакой дополнительной не нужно, ибо в Контакте практически весь рунет зарегистрирован. Вот только по поводу сохранности созданных коротких ссылок сказать ничего не могу, ибо все зависит от текущей политики ВК. - Кликер от Яндекса — еще один довольно-таки простенький сервис (но от именитого разработчика, что дает надежду на стабильность его работы), позволяющий сократить ссылку. Опять же имеется только форма для ввода целевого Урла и кнопка. Правда тут после окончания процесса укорачивания еще и картинку с QR-кодом вам предлагают скопировать, в отличии от рассмотренного выше сократителя ссылок от ВК. Вполне может пригодиться.
На мой взгляд не хватает функциональности, но зато надежность должна быть на высоте в силу серьезности бренда приютившего этот сокращатель на своих мощностях.
- Bitly.com — раньше этот укорачиватель размещался по адресу bit.ly, если кто помнит. Для сокращения адреса любой страницы нужно будет лишь вставить ее в приведенное поле и нажать на кнопку «Shorten». Будет проведено мгновенное сокращение и вам предложат скопировать полученный в результате укорот:
Все сделанные вами за данную сессию короткие ссылки будут отображаться ниже (по ним даже статистику переходов можно будет посмотреть), и это все без регистрации (видимо, вас идентифицируют по кукам сохраненным в браузере).
Однако, если вы пройдете регистрацию в сокращателе bit.ly (для этого можно использовать уже существующие аккаунты в соцсетях), то сможете получить доступ к расширенной статистике, а также сможете изменять вид полученных коротких ссылок по своему усмотрению (если такой вариант уже кем-то не используется):
Укороченные линки начинающиеся с j.mp. Ненужные ссылки можно будет отправлять в архив, а также стоит знать, что для переадресации сервис использует 301-редирект, что дает определенную надежду на то, что по такой ссылке передается вес (это важно владельцам сайтов и СЕО-шникам).
- TinyUrl.com — один из первых сервисов-сокращетелей, который за это время набрал довольно-таки солидную армию приверженцев. Правда его текущий внешний вид несколько не соответствует взыскательным требования пользователей, но работает он вполне исправно.
Как видно на скриншоте, при желании вы можете поменять предлагаемый Урл на что-то осмысленное. Какого-то еще дополнительного функционала данный сервис не предоставляет.
- U.TO — данный сервис-укоротитель принадлежит компании uCoz. Ссылка в результате получается еще короче, чем у большинства конкурентов, в силу очень короткого доменного имени сервиса. Если вы пройдете регистрацию, то получите доступ к статистике кликов по укороту, а также аудитории их производящей (страна, браузер, ОС и т.п.).
 Да и регистрации никакой дополнительной не нужно, ибо в Контакте практически весь рунет зарегистрирован. Вот только по поводу сохранности созданных коротких ссылок сказать ничего не могу, ибо все зависит от текущей политики ВК.
Да и регистрации никакой дополнительной не нужно, ибо в Контакте практически весь рунет зарегистрирован. Вот только по поводу сохранности созданных коротких ссылок сказать ничего не могу, ибо все зависит от текущей политики ВК. Будет проведено мгновенное сокращение и вам предложат скопировать полученный в результате укорот:
Будет проведено мгновенное сокращение и вам предложат скопировать полученный в результате укорот: Правда его текущий внешний вид несколько не соответствует взыскательным требования пользователей, но работает он вполне исправно.
Правда его текущий внешний вид несколько не соответствует взыскательным требования пользователей, но работает он вполне исправно.Как можно заработать на коротких ссылках?
Как ни странно, но на этом деле можно и заработать. Существуют специальные сервисы, которые позволяют не просто сократить ссылки, но и получать деньги за переходы по ним. Дело в том, что на промежуточной странице пользователю будет показываться небольшая реклама, на которой этот сервис и зарабатывает, делясь частью прибыли с вами. Получается что-то вроде рекламы при переключении каналов спутникового телевидения (типа Триколора).
Получается что-то вроде рекламы при переключении каналов спутникового телевидения (типа Триколора).
Причем такие укороченные ссылки можно размещать как у себя на сайте, так и в любом другом месте интернета (форумах, соцсетях, блогах и т.д.). Любой переход по ним будет оплачиваться, что вкупе может вылиться во вполне приличные суммы при должном подходе. В принципе, подобных сервисов много, но из них я бы выбрал один проверенный, которому можно доверять:
- CatCut — здесь одна из самых высоких ставок за переход по распространяемой вами ссылке. Создаются укороты прямо в интерфейсе сервиса (нужно будет на главной странице выбрать вариант заработка «На своих ссылках»).
Вставляете оригинал (например, http://ktonanovenkogo.ru), получаете укроченную (http://catcut.net/7LL7) и именно ее вставляете на форуме, соцсети и в других местах.Не забудьте в настройках (вкладка «Заработок на ссылках») включить показ рекламы, при переходе по укороченным в сервисе ссылкам (иначе доход они приносить не будут):
Так же возможны и другие варианты заработка, о чем можно почитать у них на сайте.
Вывод денег возможен на Вебмани и Яндекс Деньги. Минимальная сумма для вывода один рубль, что есть гуд.
 Вывод денег возможен на Вебмани и Яндекс Деньги. Минимальная сумма для вывода один рубль, что есть гуд.
Вывод денег возможен на Вебмани и Яндекс Деньги. Минимальная сумма для вывода один рубль, что есть гуд.Если пользуетесь еще каким-то сокращателем с возможностью заработка, то не примените поделиться своим опытом и уровнем доходов.
Какой сократитель ссылок выбрать?
Тут все зависит от стоящих перед вами задач.
- Если вам требуется укоротить Урл какой-то страницы раз в месяц и потом нет необходимости отслеживать статистику переходов или просто обращаться к списку ранее ваших сокращенных адресов страниц, то чем проще будет сервис, тем лучше. Учитывать стоит, пожалуй, только их надежность, поэтому тут можно порекомендовать http://vk.com/cc или http://clck.ru/ в силу их простоты и принадлежности к крупным игрокам рунета.
- Если же вы проводите, например, планомерное сокращение ваших реферальных ссылок, то тут очень желательно будет иметь возможность периодически обращаться к их списку и просматривать статистику перехода по ним. Тут однозначно можно рекомендовать goo.gl или Bitly.com.
- Если планируете использовать короткие ссылки в Емайл-рассылках, то лучше выбрать Bitly.com, ибо Гоо.гл, как уже упоминал, может вдруг перестать открывать укороты под предлогом подозрения в рассылке спама.
- Если кроме всего прочего вы хотите еще и заработать на укораченных ссылках, то можете попробовать CatCut
 Тут однозначно можно рекомендовать goo.gl или Bitly.com.
Тут однозначно можно рекомендовать goo.gl или Bitly.com.А в остальном все определяется вашим вкусом, предпочтениями или привычками.
Удачи вам!
Как сократить ссылку: обзор 11 сервисов
Источник: texterra.ruАвтор: Мария Журавлева
Сокращатели URL – это сервисы, которые позволяют преобразовать длинные адреса в более короткие и удобные. 30 марта 2019 Google окончательно закрыл один из самых популярных – Google URL Shortener (goo.gl). Однако остались еще 11 инструментов, с помощью которых можно сокращать ссылку. Разбираем их функциональность и тарифы.1. is.gd
Is.gd – англоязычный бесплатный сокращатель ссылок от сайта Memset.Результат сокращения: внизу ссылка на статистику
Длинную ссылку вида http://texterra.ru/blog/pochemu-kopirayteram-pora-perestat-slushat-marketologov-i-poyti-uchitsya-pisatelskomu-masterstvu.html он легко сокращает до https://is.gd/Nh4ywp. Чтобы отследить статистику, при создании ссылки нажимаем на «Further options» и ставим галочку «Log statistics for this link».
Включаем статистику
В статистике можно посмотреть число переходов по ссылке и отсортировать их по дате, стране, браузеру и т. д. Для просмотра статистики после создания ссылки нажимаем на «I want to see statistics for this URL».
Результат сокращения: внизу ссылка на статистику
Статистика кликов по ссылке
История созданных ранее ссылок доступна без регистрации и сохраняется благодаря данным cookies. Сервис работает стабильно, предоставляет статистику, но русскоязычным пользователям может быть не совсем удобным.
2.
 Bit.do Сервис bit.do был создан бразильской интернет-компанией Insite. Сами разработчики выделяют следующие преимущества:
Bit.do Сервис bit.do был создан бразильской интернет-компанией Insite. Сами разработчики выделяют следующие преимущества:
-
Возможность регистрации собственного домена, например, yourshortna.me. Базовый тариф обойдется в 85 долларов в месяц, а Enterprise будет стоить 250 долларов.
-
Статистика по популярности и кликабельности вашей ссылки, в том числе по отдельным странам и городам.
-
Возможность задать короткое имя ссылки.
-
Созданные ссылки навсегда останутся рабочими.
Стартовая страница сервиса
Испытаем сервис: вставляем в строку ссылку, которую хотим сократить – http://texterra.ru/blog/kak-proanalizirovat-dostizhenie-tseley-v-google-analytics-pri-nepravilnykh-nastroykakh.html, задаем ей сокращенное имя, ставим галочку антикапчи, нажимаем кнопку «Shorten».
Вводим ссылку и короткое имя для нее
Получаем две ссылки и QR-код:
Результат сокращения
Первая ссылка – укороченная версия исходной: http://bit. do/GoogleAnalytics. Другая показывает аналитику:
do/GoogleAnalytics. Другая показывает аналитику:
Статистика показывает число просмотров, географию пользователей и другие данные
Сервис довольно простой и удобный, главные плюсы – подробная аналитика и возможность задать собственное имя ссылки.
UPD Короткое имя, которое задает пользователь, должно быть уникальным — никогда ранее не использоваться в сервисе. Если такую комбинацию символов вводил кто-либо до вас, сервис выдаст ошибку, придется придумывать новое.
3. Bitly.com
Bitly – один из самых популярных сервисов для сокращения URL.
Главная страница сервиса
Bitly не просто сокращает ссылки, но и предоставляет статистику переходов по ним. Для ее получения нужно зарегистрироваться или зайти через Twitter или Facebook. Или просто добавить знак плюса к сокращенному URL, вставить в адресную строку браузера и перейти.
Пример аналитики по созданной ссылке
При создании ссылки можем дать ей короткое имя, для этого вводим его в разделе «Customize». Сервис уверяет: кастомные ссылки дают на 34 % больше кликов.
Сервис уверяет: кастомные ссылки дают на 34 % больше кликов.
Пример короткого имени ссылки
Существует и платная версия – ее стоимость начинается от 29 $ в месяц и зависит от количества обычных и пользовательских ссылок, продолжительности их действия и сохранения отчетности.
По моему мнению, это отличное, стабильно работающее приложение с множеством функций.
UPD Некоторые провайдеры в России блокируют ссылки, сокращенные через Bitly. У людей, которые пользуются услугами таких провайдеров, ссылки не открываются.
4. Tiny URL.com
Tiny URL был запущен в 2002 году, это один старейших и популярнейших сервисов по сокращению ссылок. С ним какое-то время работал «Твиттер», пока сначала не перешел на bit.ly, а затем не разработал собственную автоматическую систему t.co.Стартовая страница
Для начала вводим адрес ссылки. В поле «Custom alias» вы можете изменить URL на уникальный. Для этого нужно указать название, которое хотим дать ссылке.
Сервис довольно удобный и простой, хорошо справляется со своей задачей.
5. U.to
U.to – русскоязычный сервис по сокращению ссылок.
Внешний вид сайта
Для получения подробной статистики на сайте необходимо зарегистрироваться, но для сокращения ссылки регистрация необязательна.
Из ссылки http://texterra.ru/blog/obraz-geroya-kak-izobrazheniya-uvelichat-vashu-konversiyu-12-primerov.html делаем http://u.to/YgdIDw, нажав кнопку «Сократить».
Сокращенная ссылка
Для просмотра статистики по ссылке нужно нажать на соответствующую кнопку.
Так отображается статистика кликов по ссылке
В целом, сервис довольно простой, хотя устаревший. Особенно бесит форма регистрации, она у него самая долгая по сравнению с другими.
6. Cutt.us
У сервиса Cutt.us существует две версии.Первая – стандартный сокращатель ссылок. Вставляем http://texterra.ru/blog/rekruting-kak-marketing-ili-konvertiruem-soiskateley-v-pokupateley. html → задаем уникальное имя ссылки в строке «Suffix» → нажимаем «Cut».
html → задаем уникальное имя ссылки в строке «Suffix» → нажимаем «Cut».
Главная страница сервиса с введенной ссылкой
Получаем короткую ссылку http://cutt.us/recruit и QR-код.
Готовая ссылка
Вторая версия сервиса Smart Multi URL Shortener предназначена для обработки сразу нескольких длинных ссылок. Вводим три URL и задаем уникальное имя «helloblog». В итоге получается:
Вставляем необходимые ссылки
Нажимаем «Gooo..!» и получаем несколько сокращенных ссылок.
Получаем сокращенные ссылки, объединенные одним словом
Главная особенность сервиса – возможность создания сразу нескольких ссылок, что довольно удобно, хотя сервис показался более медленным, чем остальные в обзоре.
7. Vk.com/cc
Vk.cc – официальное приложение VK для сокращения ссылок. Доступно только зарегистрированным пользователям «ВКонтакте», однако созданные в нем ссылки можно использовать и вне этой соцсети.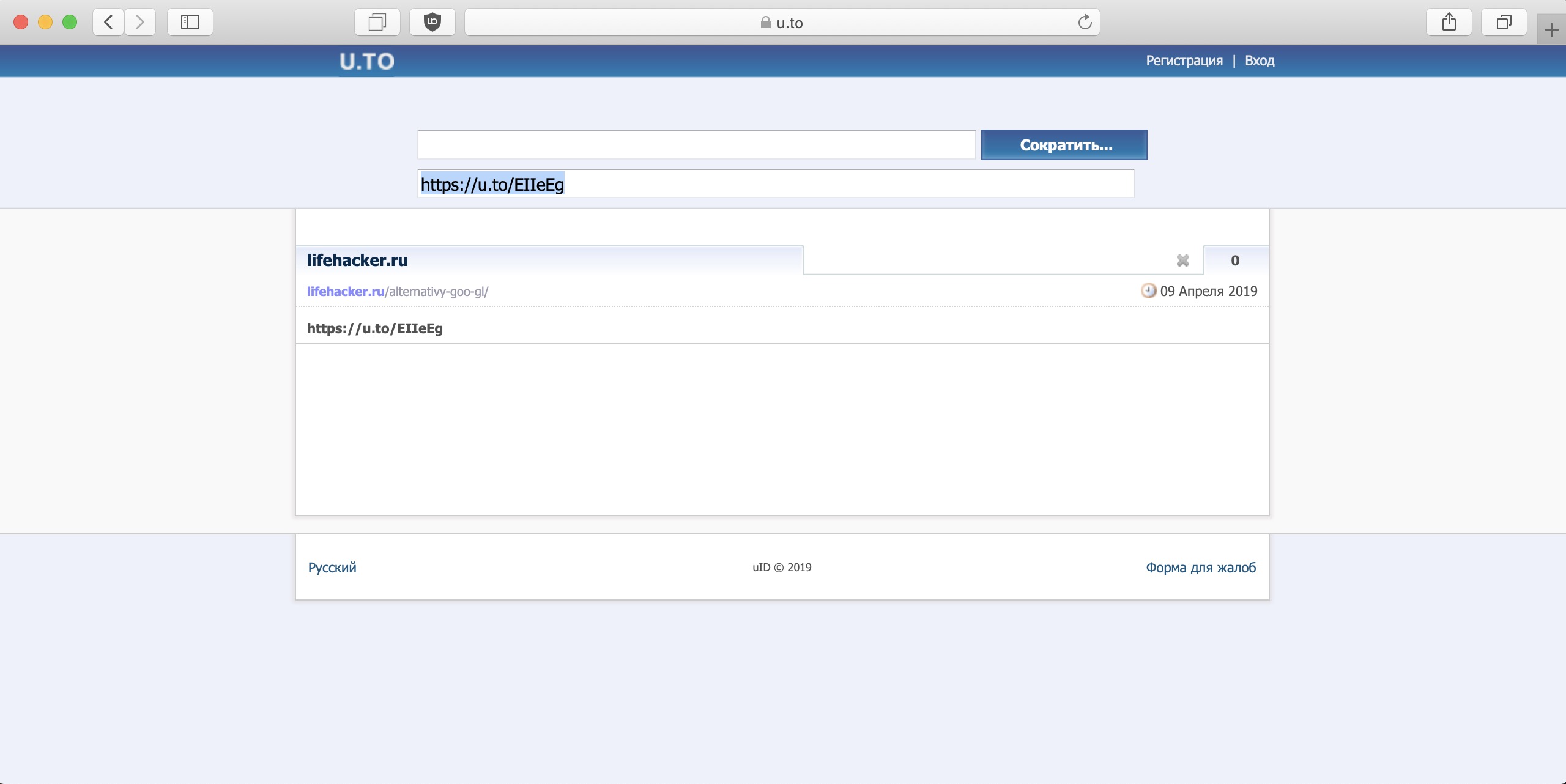
Так выглядит инструмент vk.cc
Кстати, эту статистику можно закрыть от посторонних, если поставить соответствующую галочку до сокращения ссылки. Тогда URL в списке будет помечено замочком.
Фрагмент подробной статистики по сокращенной ссылке
Приложение нравится своей простотой и стабильностью работы.
8. Clck.ru
В 2010 году «Яндекс» запустил свой собственный сервис по сокращению ссылок – «Кликер».Сервис русскоязычный и достаточно простой.
Главная страница сайта
«Для настоящих гиков» сайт предлагает следующее:
Страница с кодом
Помимо ссылки, сервис создает и QR-код, а также загадочную инструкцию. Каждый символ ссылки дан в русской транскрипции, видимо, для удобства диктования его по телефону, хотя функция выглядит довольно странно. Полученную ссылку можно сразу расшарить в соцсетях при помощи кнопки «Поделиться».
Полученную ссылку можно сразу расшарить в соцсетях при помощи кнопки «Поделиться».
Итоговый результат
Сервис хорошо и стабильно работает, неудобство заключается только в отсутствии аналитики.
9. Lnnkin
Lnnkin – еще один зарубежный сокращатель ссылок. Можно задать кастомный URL вида lnnk.in/@shlink, плюс даже защитить ссылку паролем.
Главная страница сервиса
Чтобы пользоваться функционалом сервиса, придется зарегистрироваться. Без этого сократить даже одну ссылку без возможности отслеживать клики не получится.
Поле регистрации попадаем в личный кабинет с формой генерации шортлинков и опциями управления ссылками.
Простая форма сокращения ссылок
Чтобы проанализировать статистику переходов, перейдите в раздел Manage Links личного кабинета и кликните соответствующую иконку рядом с нужной ссылкой.
Открываем статистику по сокращенной ссылке
Здесь же ссылку можно отредактировать, отключить, отправить в архив. Некоторые функции доступны только на платных тарифах.
Статистика по ссылке выглядит так:
Ниже можно посмотреть статистику по гео, ОС, браузерам, устройствам
Есть платная подписка для тех, кому нужно, например, редактировать более 5 ссылок или быстро создавать через API свыше 5 000 ссылок в месяц. Но в целом хватит бесплатных возможностей, ведь нет ограничений на количество обычных коротких и пользовательских URL, все ссылки с пожизненным сроком действия.
Кстати, у Lnnkin есть еще расширение для Chrome, позволяющее сокращать ссылки прямо из вкладки браузера.
Так работает расширение для «Хрома»
Получается, это простой, практически бесплатный и в то же время многофункциональный инструмент. Однозначно 10 из 10.
10. Rebrandly
Rebrandly помогает превратить «длинные, уродливые и непроизносимые» URL-адреса в кастомные фирменные ссылки.
Чтобы сократить ссылку, нужно:
- Зарегистрироваться.
- Вставить URL на главной и нажать кнопку «Rebrand». Или кликнуть на «New link» в разделе «Links», затем указать ссылку.
- Выбрать способ сокращения: генерировать самый короткий адрес, SEO-дружественный (учитывающий слова в URL) или, например, абсолютно случайный.
- Указать домен – оставить rebrand.ly или прикрепить собственный.
- Добавить к исходной ссылке UTM-метки, если это необходимо.
 Или кликнуть на «New link» в разделе «Links», затем указать ссылку.
Или кликнуть на «New link» в разделе «Links», затем указать ссылку.Так работает сокращение ссылок в Rebrandly: здесь выбрали SEO-дружественный URL, оставили собственный домен сервиса, UTM не добавили
Бесплатная статистика ограничена количеством кликов. Чтобы проводить подробную аналитику (следить за географией, устройствами, языками), а также подключать несколько доменных имен, постить сокращенные ссылки тысячами, строить отчеты – придется платить. Минимальный тариф стоит 29 $ в месяц.
Что тут сказать: крутой профессиональный инструмент, который особенно пригодится крупным брендам и медиа. Очень удобно, что сервис позволяет не отходя от кассы генерировать UTM-метки, регистрировать домены и даже собирать аудитории для ретаргетинга.
11. To.click
To.click – мощный инструмент для создания ссылок трех типов: простые сокращенные, таргетированные (с пикселем, например, от «Фейсбука») и диплинки (для перехода на конкретную страницу внутри приложения). По умолчанию URL начинается с clc.to, но также можно подключить свой домен.Так же, как и в Lnnkin, чтобы пользоваться To.click, придется зарегистрироваться. А еще — подтвердить свой email. Без этого ни попасть в личный кабинет, ни сократить ссылку не получится.
После регистрации попадаем в панель управления. Чтобы открыть форму сокращения ссылок, кликаем соответствующую ссылку в правом верхнем углу.
В форме задаем короткое имя, вставляем ссылку, выбираем пиксели (если добавили их ранее через соответствующий раздел левого меню) и при необходимости ставим галочку «Закрыть статистику от всех».
Также можно выбрать домен из выпадающего списка — варианты от To.click или свой, если вы его привязывали
Плюс есть API и телеграм-бот, скоро обещают интеграцию со Slack. А в панели управления можно отследить клики, источники переходов, географию пользователей, их платформы и языки.
А в панели управления можно отследить клики, источники переходов, географию пользователей, их платформы и языки.
Фрагмент статистики по ссылкам
***
Кстати, вот еще. Если пользуетесь Zapier, там есть встроенный URL Shortener. В Hootsuite тоже свой, называется Ow.ly. А если разбираетесь в программировании, можно запустить собственный инструмент, используя Polr и Yourls.зачем, что умеют, как использовать
Инструменты сокращения ссылок нужны, чтобы сократить длинные URL для использования в социальных сетях, презентациях и других документах. Для вас мы подготовили обзор лучших сервисов, которые помогают сократить ссылку, поделиться ей, создать QR код.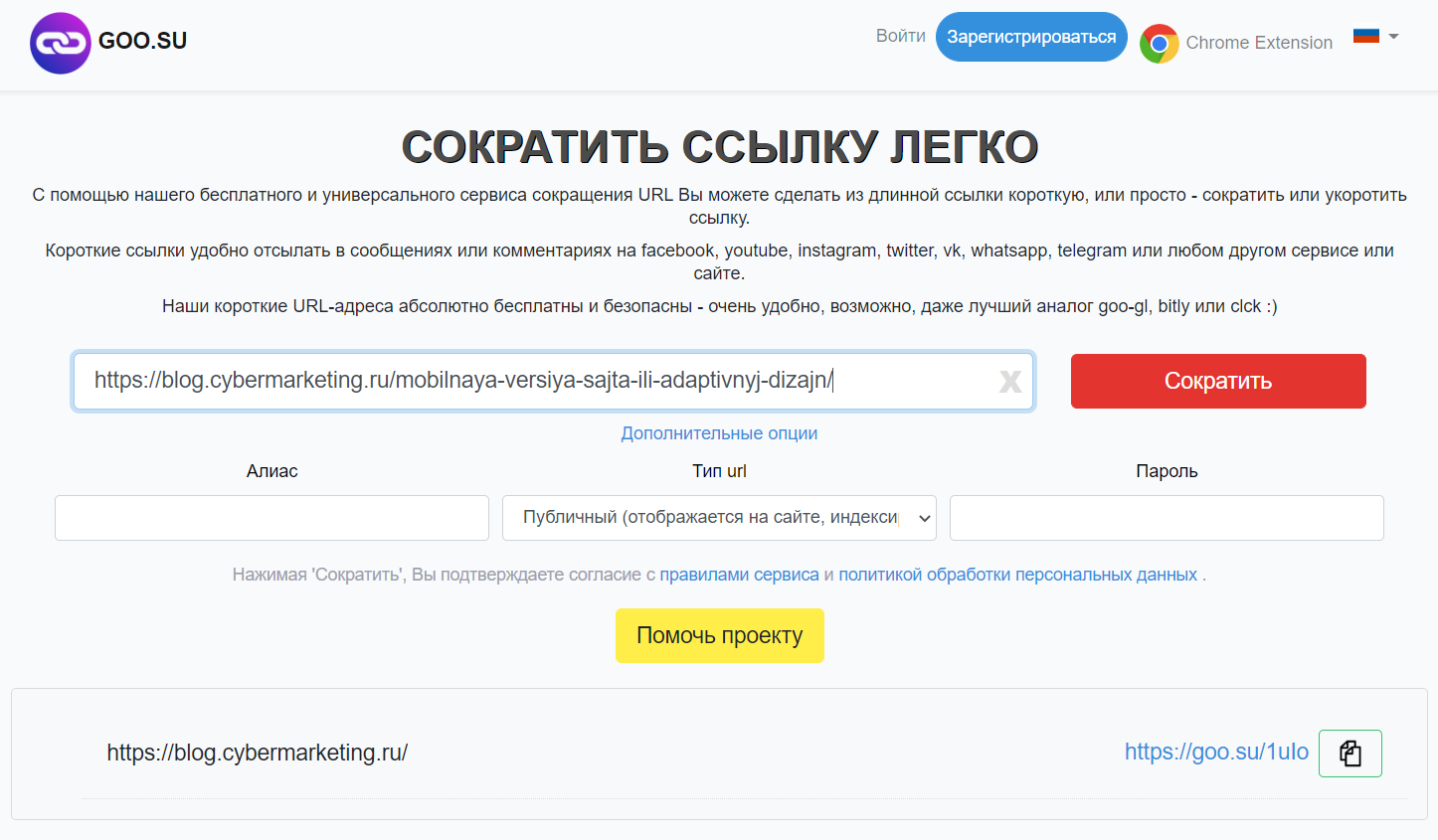
Почему ссылки при передаче приходится укорачивать?
Почему возникла потребность в сервисах для такой мелкой задачи? Ссылки сокращают по двум основным причинам:
-
Короткие URL аккуратнее смотрятся в отправляемых сообщениях, постах и презентациях, текст с такими ссылками проще форматировать.
-
После сжатия URL будет проще продиктовать и запомнить. При этом не потеряются UTM метки. Даже если адрес наберут с распечатки, вы сможете это понять!
Сервисы сокращения URL также служат аналитическим инструментом. Многие из них показывают статистику переходов по ссылке с достаточно подробной аналитикой по географии, каналам трафика.
Всегда ли уместно и правильно укорачивать длинные ссылки?
Использовать сокращение ссылок не всегда правильно и удобно. Например, в e-mail письма с сокращенными ссылками попадут в спам. Это мера предосторожности, ведь спрятанные под сокращения URL могут вести на вредоносные сайты или к файлам с вредоносным ПО.
Например, в e-mail письма с сокращенными ссылками попадут в спам. Это мера предосторожности, ведь спрятанные под сокращения URL могут вести на вредоносные сайты или к файлам с вредоносным ПО.
Чтобы не попасть в спам, используйте в своих рассылках полные линки. Из них можно удалить параметры, это сделает URL короче.
В навигации по сайту ссылки тоже рекомендуем не сокращать. Во-первых, здесь их никто не видит, во-вторых, ссылка может неожиданно перестать работать. Если это произойдет, сокращенные URL тоже перестанут быть активны.
Лучшие сервисы для форматирования ссылок и добавления меток
Инструменты, которые мы перечислим в подборке, полезны не только для форматирования адресов. Они позволяют добавить метки к ссылкам. А это важно при настройке маркетинговой аналитики и отслеживании того, как работает каждый из каналов продвижения.
Бизнес-инструментарий от создателей Bit.do
Максимально простой и интуитивно понятный сервис. Последнюю часть ссылки вы сможете прописать сами, это очень важно, потому что если в URL будет непонятный набор символов, пользователи не захотят по нему переходить.
Последнюю часть ссылки вы сможете прописать сами, это очень важно, потому что если в URL будет непонятный набор символов, пользователи не захотят по нему переходить.
Вместе с короткой ссылкой вы получите и соответствующий QR код, он приведет пользователя по тому же адресу на сайте. Коды можно распечатывать, использовать для оффлайн рекламы, визиток, баннеров.
Чтобы посмотреть статистику URL, нужно вставить адрес в строку браузера и добавить символ — .
Сервис разработала бразильская компания Insite. Разработчики акцентируют внимание на нескольких преимуществах:
-
Возможность зарегистрировать на сервисе собственный домен. Правда, базовый тариф обойдется недешево — в 85 USD. Enterprise тариф составляет 250 USD.
-
В статистике по всем вашим переформатированным ссылкам доступна детализация по странам и даже городам.
-
Ссылки навсегда остаются рабочими, даже если вы удалите аккаунт.

Сервис сразу генерирует уменьшенную ссылку, отдельный адрес для аналитики переходов и QR код. Важный совет! Чтобы сервис работал корректно, задавайте уникальные имена ссылок.
Из недостатков сервиса можно назвать несколько устаревший дизайн и неудобный формат отображения статистики.
Bit.ly — один из лидеров в конкурентной гонке
Один из самых популярных, удобных и функциональных сервисов. Его можно использовать даже без регистрации, но с аккаунтом он будет намного удобнее. Также доступна авторизация через Facebook и Twitter.
Только зарегистрированным пользователям. Вы увидите число кликов по дням, диаграмму с разделением по источникам трафика и пр. Также Bitly покажет информацию о том, с какого канала вы получаете больше всего переходов.
Некоторые интернет-провайдеры блокируют ссылки, созданные через этот сервис. Поэтому в единичных случаях могут возникать неудобства.
Единственным минусом может показаться не русифицированный интерфейс.
Cutt.us
Бесплатный и не требующий регистрации инструмент. Интерфейс здесь тоже только на английском. Ссылки тут можно объединить под одним ключевым словом. Также есть возможность задать нужное окончание URL адреса.
Чтобы это сделать, зайдите на сайт, вставьте в окно несколько ссылок с одного сайта. Затем просто введите общее окончание для ссылок, например: myblog.
Среди недостатков отсутствие аналитики, интерфейс без перевода на русский.
Is.gd
Легкий и удобный сервис для сокращения. Здесь можно работать не создавая аккаунта. Сохраняет статистику переходов на основе данных из cookies. Правда, интерфейс сервиса совсем не юзер френдли.
Ok.lt — для тех, кто использует Однокласники
Сервис сокращает ссылки в рекламных постах сети Одноклассники. Не рекомендуем использовать на других площадках, чтобы не запутать пользователей. Есть удобная и подробная статистика переходов по каждому из адресов. Не будет работать для украинских пользователей.
Lnnkin.com
Имеет стандартный набор функций: кастомизация окончаний в ссылках, аналитика. Также созданные ссылки можно защитить паролями. Из недостатков можно выделить большое количество всплывающих окон и рекламы на сервисе.
Отслеживать статистику по ссылкам вы можете даже без регистрации. Для этого просто сохраните в закладках страницу статистики.
У сервиса есть собственное расширение для Google Chrome, которое позволяет создавать сокращенные ссылки прямо из браузера.
Еще один недостаток в том, что редактировать короткие ссылки можно только в платной версии.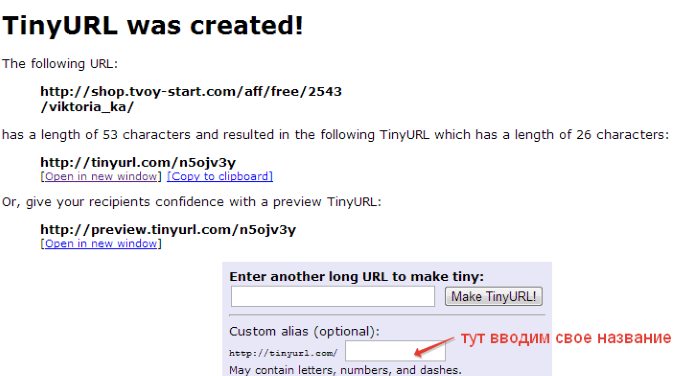
Goo.su — shortener с собственным плагином для браузера
Легкий в использовании сервис. Полученные через сократитель ссылки при необходимости можно закрыть от индексации. Эту функцию можно использовать если вам нужно разместить URL стороннего ресурса на вашем сайте. Также можно легко создать кастомный короткий адрес ссылки.
У этого сайта тоже есть свое расширение, работающее в Chrome.
Аналитика будет доступна после регистрации, а на самом сайте очень много рекламы. Это иногда причиняет неудобства.
Rebrandly.com — сервис для сокращения и продвинутой аналитики
Один из самых удобных и функциональных инструментов в нашей подборке. Этот сайт может:
-
прикрепить к ссылке другие домены, кроме шаблонного, эта функция доступна только после того, как вы приобретете платный тариф;
-
добавить к исходным ссылкам utm метки для отслеживания переходов;
-
генерировать несколько вариантов сокращенной ссылки.
В бесплатном тарифе есть ограничение на количество кликов по ссылкам. Максимальный тариф на сервисе: 29 USD ежемесячно.
Самый серьезный недостаток — в бесплатной версии недоступна глубокая аналитика.
Tinyurl.com — старый, проверенный, доступный
Один из самых старых бесплатных сервисов сокращения. Запустили его еще в 2002 году. Длительное время сайт плотно сотрудничал с Twitter. Но социальная сеть сначала переориентировалась на Bitly, а потом и разработала собственный инструмент t.co. Он работает надежно, но переполнен рекламой. Здесь тоже можно сделать уникальный URL с вашей собственной структурой и окончанием. Для этого добавьте длинную ссылку и перейдите в поле custom alias. Там формируется кастомное окончание URL.
Интерфейс здесь устаревший, но довольно понятный.
To.click — инструмент с собственным Telegram ботом
Это многофункциональный и очень продвинутый сервис. Он выходит за рамки просто сократителя ссылок. После регистрации вы сможете кастомизировать короткий URL под домен вашего сайта.Также вам будет доступен пиксель таргетированной рекламы и другие инструменты аналитики.
Он выходит за рамки просто сократителя ссылок. После регистрации вы сможете кастомизировать короткий URL под домен вашего сайта.Также вам будет доступен пиксель таргетированной рекламы и другие инструменты аналитики.
Также можно сокращать ссылки через бот @toclick_bot.
Интересная особенность в том, что сервис поддерживает даже кириллицу и эмодзи в окончании URL. Это повышает кликабельность ссылок.
Из недостатков выделим то, что полный набор возможностей доступен только после оплаты любого из тарифов.
U.to — минималистичный и предельно простой
Сервис с минималистичным дизайном. Для начала работы регистрироваться не обязательно. Но если вы захотите отслеживать переходы по ссылкам, завести аккаунт нужно.
Минусом можно назвать только слишком подробную форму регистрации с десятью обязательными полями.
Vk.com/cc — собственная разработка ВКонтакте
Сервис от социальной сети ВКонтакте. Можно использовать и за пределами сайта. Для получения сокращенных ссылок обязательно залогиниться в сети ВКонтакте. Вместе со ссылкой сервис генерирует QR код в форматах PNG и SVG.
Можно использовать и за пределами сайта. Для получения сокращенных ссылок обязательно залогиниться в сети ВКонтакте. Вместе со ссылкой сервис генерирует QR код в форматах PNG и SVG.
Также есть детальная статистика просмотров.
T1p.de — немецкая надежность в работе с линками
Фишка этого сервиса в повышенной безопасности, заботе о безопасности, сохранности личных данных. Все ссылки проверяются на защищенность и отсутствие вирусов. Сервис не хранит личные данные пользователей. Есть также функция анонимизации при переходе по ссылкам.
Минусами можно назвать не слишком интуитивный интерфейс, отсутствие кастомизации.
Сервисы, которые лучше не использовать
Есть несколько сервисов, которые мы по разным причинам советуем не использовать. Возможно, в дальнейшем их улучшат, но на момент написания статьи эти сайты по разным причинам нам не понравились:
Bc. vc — при нажатии на сокращенный адрес вы сначала видите навязчивую стороннюю рекламу. Это отнимает время и отвлекает от целей перехода.
vc — при нажатии на сокращенный адрес вы сначала видите навязчивую стороннюю рекламу. Это отнимает время и отвлекает от целей перехода.
Demo.polr.me — неплохой, но пока слишком сырой проект. Все еще в стадии бета-версии. Здесь уже есть возможность кастомизации ссылок и другие функции. Пока этот сервис вызывает впечатление ненадежного и мы не можем его рекомендовать.
Zapier URL Shortener — удобный и надежный сервис с хорошим функционалом. Но он доступен только тем, кто зарегистрировался в сервисе интеграций Zapier. Для многих это неудобно.
Yourls — это не онлайн сервис, а набор php скриптов. С его помощью вы можете создать собственный сервис сокращения ссылок. Но такой подход не всегда оправдан. Чтобы настроить и запустить, нужно привлекать разработчиков.
Сервисы Vk-cc.ru и Urlways.com мы не рекомендуем из-за того, что при открытии сокращенных ссылок пользователям приходится просматривать рекламу. Это нужно для существования сервисов, но пользователям не всегда приятно отвлекаться на просмотр.
Какой сервис использовать?
Функционал большинства сервисов очень похож. Но мелкие детали создают серьезную разницу. Нельзя назвать один сайт однозначно лучшим. Поэтому мы выделили несколько ситуаций, в каждой из которых рекомендуем использовать разные сервисы:
- Нужна просто сокращенная ссылка без дополнительных функций? Подойдут U.to, Bitly.com, Goo.su, ls.gd.
- Хотите сгенерировать QR-код для лендинга, флаера, наружной рекламы или других потребностей? Используйте Bit.do, Сlck.ru или Vk.com/cc.
- Для сокращения нескольких ссылок сразу подойдет Cutt.us.
- Для тех, кому нужен продвинутый инструмент с кастомизацией и другими дополнительными возможностями, подойдет Rebrandly.com.
Общие советы по сокращению URL
Продвинутые пользователи очень внимательно относятся к ссылкам. Они присматриваются, прежде чем переходить по ним. Также существуют нормы digital этикета, которые следует соблюдать при сокращении URL и обмене ими.
Также существуют нормы digital этикета, которые следует соблюдать при сокращении URL и обмене ими.
Есть несколько советов, которые помогут вам избежать неудобств при работе со ссылками:
-
Если вы решили, что сократить URL будет уместно, используйте сервис с https соединением, это важно для безопасности данных и доверия других пользователей к ссылкам;
-
Обязательно поставьте перед ссылкой комментарий о том, что скрывается под сокращением, по ссылкам без объяснения переходят неохотно, опасаясь вирусов;
-
Настраивайте кастомные окончания. Сделайте их понятными для человека. Например: uslugi_kompanii, kp_kompanii и пр. Особенно это актуально, если нужно сокращать ссылки на Google Docs, видео в Youtube.
Выводы
Инструменты, сжимающие URL, помогают отслеживать переходы по ссылкам. Правильно оформленные сокращенные ссылки привлекают больше внимания в социальных сетях и на сайтах.
Сравнительно недавно к стандартным функциям сократителей добавилась еще и генерация QR-кодов. Это очень удобно и расширяет сферу применения таких сервисов. Коды нужны для упаковок продукции, рекламной полиграфии и еще десятков задач. Поэтому и сайты из нашей подборки просто must have в арсенале маркетолога и других специалистов, работающих в бизнес-сфере.
Мы рекомендуем завести аккаунты на нескольких сервисах, протестировать все и выбрать 2-3 самых удобных именно для вас.
Сокращение ссылок в контакте
Одна короткая ссылка — бесконечные возможности
Короткие ссылки, это мощный инструмент маркетолога, особенно, когда он правильно используется. Наш сервис коротких ссылок позволяет бесплатно использовать свои домены для сокращения, собирать статистику, аудитории ретаргетинга, делать сложные редиректы в зависимости от условий: местоположения, устройства пользователя и других. Создавать собственные посадочные страницы или показывать баннеры поверх исходных сайтов, ссылки на которые вы сокращаете.
Создавать собственные посадочные страницы или показывать баннеры поверх исходных сайтов, ссылки на которые вы сокращаете.
Разделяйте пользователей и показывайте им уникальный контент, например, раздельные ссылки на Android и iOs
Share your links to your network and measure data to optimize your marketing campaign’s performance. Reach an audience that fits your needs.
Вы можете подключить к системе собственный домен и создавать брендированные ссылки, как t.me или t.co со своим доменом.
Вы можете создавать собственные посадочные страницы, показывать баннеры поверх других сайтов или собирать статистику их посещаемости. Создайте рекламное предложение, собственную страницу переадресации или просто установите отслеживающие пиксели или код google analitics на страницу и показывайте сайт назначения в iFrame
Для перехода на любую страницу в интернете требуется ссылка. Ее вид зависит от домена сайта и количества вложений разделов, ведущих к размещенному материалу. От этих параметров изменяется общая длина ссылки, которая в некоторых случаях может быть весьма большой. Чтобы представить адрес в укороченном виде используется сервис сокращения ссылок ВК. В текущей статье разберемся в необходимости сокращения количества символов адреса страницы, опишем процедуру использования соответствующего сервиса социальной сети и предоставим несколько сторонних ресурсов, выполняющих аналогичные функции.
От этих параметров изменяется общая длина ссылки, которая в некоторых случаях может быть весьма большой. Чтобы представить адрес в укороченном виде используется сервис сокращения ссылок ВК. В текущей статье разберемся в необходимости сокращения количества символов адреса страницы, опишем процедуру использования соответствующего сервиса социальной сети и предоставим несколько сторонних ресурсов, выполняющих аналогичные функции.
Причины необходимости сокращения ссылки
Укорачивается адрес страницы по нескольким причинам:
- Некоторые социальные сети (например, Twitter) имеют ограничение длины публикуемой записи, и если ссылка занимает почти весь допустимый объем, то на сопутствующий текст попросту не останется места.
- Сокращенный адрес страницы выглядит более привлекательно, не вызывает недоверия со стороны пользователя.
- В укороченном адресе не получится убрать некоторые элементы, что особенно важно для участников партнерских программ.
- Обход ограничений в отношении публикуемых ссылок. Ярким примером служит видеохостинг YouTube, где в аннотациях к видеозаписи можно публиковать адрес только подтвержденного сайта.
- Система борьбы со спамом ВКонтакте с большой долей вероятности будет отображать пометку «Ссылка ведет на подозрительный сайт», если в записи разместить не измененную ссылку рекламируемого продукта.
 Ярким примером служит видеохостинг YouTube, где в аннотациях к видеозаписи можно публиковать адрес только подтвержденного сайта.
Ярким примером служит видеохостинг YouTube, где в аннотациях к видеозаписи можно публиковать адрес только подтвержденного сайта.Сокращение ссылки через сервис ВК
Чтобы сократить URL страницы с использование встроенного инструмента социальной сети необходимо:
- Перейти по адресу https://vk.com/cc.
- Скопировать ссылку страницы, которую требуется сократить. Для этого нужно открыть выбранный материал сайта в браузере, выделить и скопировать текст, расположенный в адресной строке.
- Вставить скопированный URL в соответствующую строку сервиса ВК. При желании поставить галочку около пункта «Статистика ссылка видна только вам». Это позволит скрыть анализ переходов от посторонних.
- Нажать кнопку «Сократить».
Теперь под строкой для вставки сокращаемых ссылок появится блок, отображающий недавно преобразованные адреса. Под каждым из них видна дата создания, а нажав значок в правом углу, получится открыть статистику переходов. В отчете будет предоставлена следующая информация:
Под каждым из них видна дата создания, а нажав значок в правом углу, получится открыть статистику переходов. В отчете будет предоставлена следующая информация:
- график уникальных посетителей и просмотров;
- пол и возраст;
- географическое расположение пользователей.
Сторонние сервисы сокращения ссылок
Если по каким-то причинам функционал соцсети не устраивает, можно воспользоваться сторонними сайтами, предназначенными для выполнения аналогичной задачи. Вот некоторые из них:
Работа со всеми указанными сервисами аналогична той, что представлена для ВКонтакте. Потребуется вставить в соответствующее поле ссылку и нажать кнопку «Сократить». Большинство сайтов предоставляет доступ к подробной статистике, но для этого придется зарегистрироваться. В некоторых случаях возможно даже изменить конечный вид укороченного адреса.
Справка. Широко используемый сервис goo.gl прекратил свою работу с 30 марта 2018 года. Все сокращенные URL остаются рабочими, но создание новых ссылок больше не возможно.
Сделав адрес страницы короче, пользователь получает более привлекательную ссылку, которая будет удобна для использования. Для достижения задуманного можно воспользоваться встроенным функционалом ВКонтакте или прибегнуть к сторонним сервисам. Почти все представленные сайты позволяют отслеживать статистику переходов по сокращенным URL, что обеспечивает возможность анализа заинтересованности пользователей к опубликованной ссылке.
Возможно вы уже сталкивались с проблемой, когда нужно отправить другу ссылку, а вконтакте блокирует ее. Или другая ситуация — вы хотите разместить ссылку в публичном доступе, а она длинная как простыня и выглядит некрасиво. В обоих случаях на помощь приходит сокращение ссылок ВК.
В этой статье мы рассмотрим 3 популярных сервиса для сокращения ссылок: vk.cc, Кликер и u.to, их плюсы и минусы. Попробуем сократить в них разные ссылки и выберем подходящий.
Содержание
Сервис коротких ссылок vk.cc
Сокращатель ссылок vk.cc доступен по 2 ссылкам vk.cc и vk.com/cc. Обе ссылки ведут на одну и ту же страницу.
Сервис сокращения ссылок vk.cc
В пустое поле нужно вставить ссылку, которую вы хотите сократить и нажать кнопку «Сократить».
После сокращения готовая короткая ссылка появится в этом же поле. Вам останется только скопировать ее и разместить там, где хотели или отправить в сообщении другу.
Через сервис коротких ссылок vk.cc можно просматривать статистику по сгенерированным ссылкам. В статистике можно увидеть количество переходов по вашей ссылке.
Чтобы посмотреть статистику нужно щелкнуть в область, отмеченную на скриншоте ниже. Если щелкнуть на саму ссылку, то произойдет переход по ней, а не в статистику.
Недавно сокращенные ссылки vk.cc
Можно посмотреть количество переходов по ссылке и не заходя в статистику, она видна в нижем левом углу строки.
Просмотр статистики
Просмотр статистики может понадобиться, если вы разместили вашу реферальную ссылку в какой-нибудь группе, например, и хотите посмотреть сколько людей по ней перешло, чтобы оценить, насколько эффективно было ваше объявление.
Важно! Если не поставить галочку «Статистика видна только Вам», то ее смогут посмотреть все желающие и тоже оценить эффективность вашей рекламы. Так что не забывайте ставить, если не хотите делиться с конкурентами информацией.
Все ранее сокращенные вами ссылки будут доступны на странице vk.cc в разделе «Недавно сокращенные ссылки», если вы будете залогинены в вашем аккаунте.
Одно но, этот сервис не будет сокращать ссылки, которые ему не нравятся.
А не нравится ему все, что связано со спамом, со сторонними сервисами накрутки, сомнительным заработком. Т.е. скорее всего реферальные ссылки сократить не получится.
Если вы получили сообщение «Невозможно сократить эту ссылку», то это значит, что ваша ссылка как раз попадает в список неугодных ВК.
Сообщение «Невозможно сократить эту ссылку» в сервисе vk.cc
Плюсы и минусы сервиса vk.cc
- Можно перейти прямо из Вконтакта
- Доступен просмотр статистики
- Не все ссылки можно сократить
- Требуется регистрация Вконтакте
Вывод: Сервис хорош для сокращения длинных ссылок, например, с utm-метками, подойдет для таргетологов и смм-менеджеров. Сокращает не все ссылки.
Сокращает не все ссылки.
Как сократить ссылки через Кликер
Еще один сокращатель ссылок принадлежит Яндексу и хорош тем, что не придирается к ссылкам и сокращает любые ссылки без разбора.
Находится по адресу clck.ru.
Интерфейс предельно прост: вставляете свою ссылку и нажимаете клик.
Статистики этот сервис не дает, но зато генерирует QR-код.
Для тех, кто не знает, поясню, что QR-код можно отсканировать через сканер мобильного телефона и получить из него ссылку, чтобы не набивать ее руками в телефоне.
Плюсы и минусы сокращателя ссылок Кликер
- Простой интерфейс
- Не требует регистрации
- Генерирует QR-код
- Сокращает все ссылки без ограничений
Сокращение ссылок через u.to
Еще один простой сервис сокращения ссылок.
Сервис хорош тем, что хранит и отдает статистику переходов по ссылке. Есть возможность зарегистрироваться и просматривать данные через личный кабинет.
Если хотите просматривать статистику, то лучше зарегистрироваться, т.к. сервис хоть и помнит ваши ссылки без регистрации, но после очистки куки все забудет.
Регистрация в u.to
Для регистрации нужно в верхнем правом углу экрана нажать ссылку Регистрация. Откроется форма ввода данных, которую нужно заполнить полностью!
Рекомендую в форме регистрации под Именем, Фамилией и Датой рождения в строках Могут видеть: Все заменить на никто. Ну незачем всем видеть персональные данные, как мне кажется.
После заполнения нажимаете на кнопку «Создать аккаунт», заходите на электронную почту, которую указали и переходите по ссылке для подтверждения аккаунта.
Все, теперь можно заходить в личный кабинет.
Через личный кабинет доступна полная статистика ссылок, возможность создания сокращенных, таргетированных ссылок и дип линков.
И самая интересная возможность, это делать сокращенные ссылки с привязкой к своем домену, если он у вас есть.
Так что пожалуй этот сервис можно признать одним из лучших и по простоте и по функционалу.
Плюсы и минусы сервиса u.to:
- Не требует обязательной регистрации;
- Можно создать личный кабинет;
- Сокращает все ссылки;
- Делает таргетированные ссылки и дип линки.
Как сократить ссылку вконтакте. Сокращение ссылок вконтакте через официальный сервис
Социальная сеть «ВКонтакте» в наши дни используется не только для общения. Многие активно зарабатывают на популярном ресурсе, пользуясь его многочисленными функциями. В этой статье вы узнаете, что такое сокращение ссылки «ВКонтакте» и для чего оно нужно.
Для чего нужно сокращать ссылки?
Предположим, вам нужно прорекламировать свой сайт в социальной сети. Однако большинство ссылок на сторонние ресурсы имеет очень длинные названия, которые в тексте объявления выглядят не очень привлекательно для потенциального клиента. Сокращение ссылки «ВКонтакте» позволит придать ей более компактную форму. Кроме того, у большинства пользователей даже не возникнет подозрений, что вы ссылаетесь на сторонний ресурс, а не на страничку в социальной сети.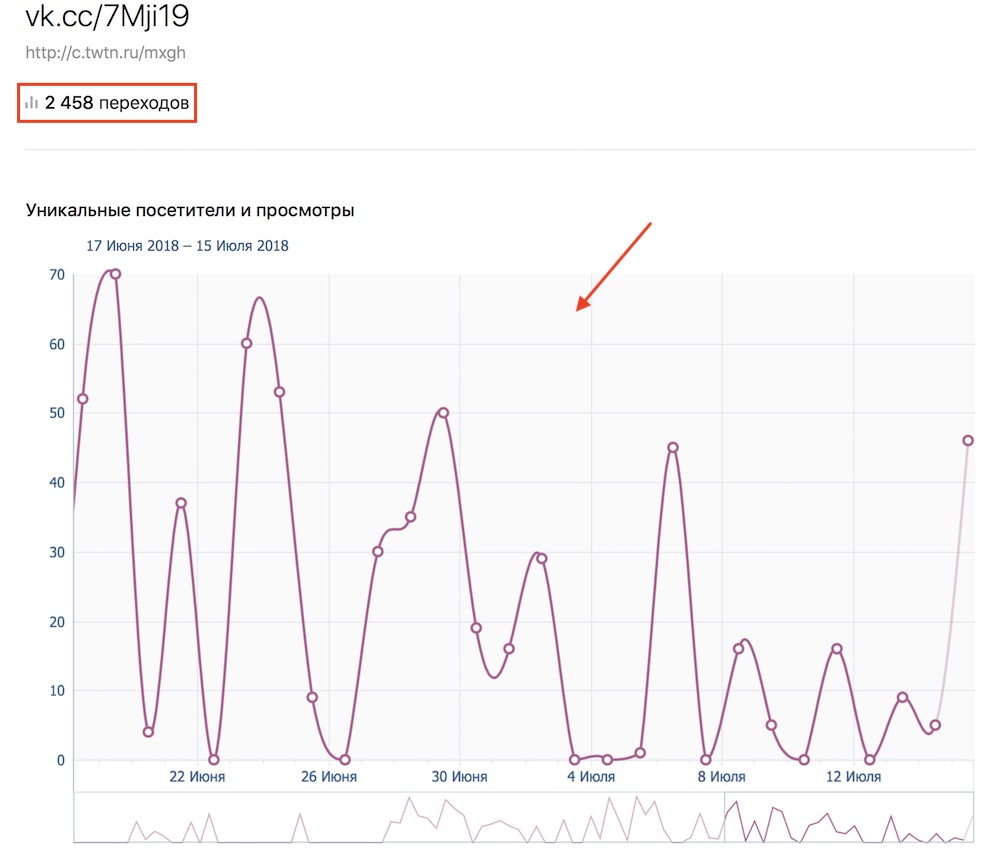
Этим активно пользуются рефереры, которые набирают большую команду рефералов для своих проектов. Однако переходы на сторонние сайты могут привести на мошеннические ресурсы. Поэтому позаботьтесь о том, чтобы на вашем компьютере стояла хорошая антивирусная программа. Ведь сокращение ссылки «ВКонтакте» не защищает вас от вредоносных сайтов. Визуально даже профессионалу невозможно определить, куда именно приведет короткий адрес, оставленный в сети.
Как сократить ссылку?
Для того чтобы сделать из длинной ссылки короткую, можно воспользоваться одним из сервисов в сети. На сегодняшний день такая услуга предоставляется совершенно бесплатно и доступна абсолютно всем. Достаточно просто скопировать в поле нужную адресную строку и система выдаст короткую ссылку за несколько секунд.
Сервис сокращения ссылок «ВКонтакте»
Эта функция в популярной социальной сети появилась совсем недавно, но уже успела принести плоды веб-мастерам. Для того чтобы воспользоваться сервисом сокращения ссылок, нужно перейти по адресу vk. cc. Страница представляет собой только после заполнения которой пользователь получит сокращенный вариант своей ссылки. Использовать полученный результат можно не только на просторах социальной сети, но и на любом сайте, который требует шифровки. При этом система не сохраняет введенные данные, что гарантирует полную конфиденциальность пользователя.
cc. Страница представляет собой только после заполнения которой пользователь получит сокращенный вариант своей ссылки. Использовать полученный результат можно не только на просторах социальной сети, но и на любом сайте, который требует шифровки. При этом система не сохраняет введенные данные, что гарантирует полную конфиденциальность пользователя.
Таким образом, сокращение ссылки «ВКонтакте» — это отличный способ быстро и надежно раскрутить свой проект и привлечь как можно больше Ведь будущее стоит за информационными технологиями. А это означает, что нужно пользоваться всеми возможностями, которые дают нам социальные сети.
В Google считают, что сокращатель ссылок отлично справляется со своей задачей, но его время прошло. Все созданные ранее в Goo.gl ссылки продолжат работать даже после закрытия сервиса.
Однако всё равно придётся искать новый инструмент для сокращения ссылок. Вот сервисы, которые хорошо справятся с этой задачей.
1. Is.gd
Бесплатный сокращатель ссылок имеет минимум возможностей и простой интерфейс. Чтобы отслеживать статистику, не надо регистрироваться, достаточно нажать на I want to see statistics for this URL под ссылкой.
Об этом сервисе Лайфхакер уже . Отличный функциональный сокращатель с быстрым редиректом и подробной статистикой.
Сокращатель ссылок от бразильской компании Insite. Сервис порадует подробной статистикой, а также возможностью задать имя для ссылки и даже создать собственный домен. За последнее, правда, придётся заплатить.
Один из ветеранов сокращательного дела. Простой и удобный сервис, который служит верой и правдой уже много лет.
5. Ow.ly
Сервис от платформы отложенного постинга Hootsuite. Чтобы воспользоваться сокращателем, придётся зарегистрироваться в сервисе. Если вы планируете пользоваться им только для сокращения ссылок, выбирайте бесплатный тариф.
6. U.to
Простой русскоязычный сервис. Чтобы посмотреть статистику, придётся зарегистрироваться. Можно обойтись и без этого, если просто хотите сократить ссылку.
Этот сервис примечателен тем, что в нём можно создавать сразу несколько ссылок.
8. Bitly
Популярный сервис с удобным интерфейсом и функциональной панелью администрирования, позволяющей отлеживать клики по ссылкам и просматривать расширенную статистику по ним.
9. Zapier URL Shortener
Отличный сервис одноименной компании для интеграции веб-приложений. Умеет не только сокращать ссылки, но и делать это автоматически при срабатывании определенных триггеров.
10. Rebrandly
Продвинутый сервис, который пригодится для продвижения бренда. С помощью Rebrandly можно создавать персонализированные ссылки с доменом вашего сайта.
11. Lnnk
Очень простой сервис сокращения, в котором можно генерировать защищенные паролем ссылки. Lnnk работает как в качестве веб-сервиса, так и в виде браузерного расширения.
12. Polr
Сервис Polr позволит быстро сократить любую ссылку и настроить ее отображение, выбрав окончание на свое усмотрение. Кроме того можно воспользоваться исходным кодом Polr и сделать собственную сокращалку.
Кроме того можно воспользоваться исходным кодом Polr и сделать собственную сокращалку.
Согласитесь, что длинный урл адрес не всегда смотрится красиво. Как это исправить?
Уже давно существует большое количество сервисов, которые сокращают ссылки. Вы указываете оригинальный урл, и формируете сокращенный вариант. При переходе по такой ссылке, пользователь попадает на нужную страницу.
https://vk.com/ccВот так выглядит главная страница.
Скопируйте нужный урл адрес, и вставьте его в форму. Затем нажмите кнопку «Сократить» .
Вставляем ее в форму, и формируем сокращенную ссылку.
Мы получим готовый урл адрес. Он выглядит вот так:
Https://vk.cc/6huY1D
Можете скопировать его, и использовать по назначению. Попробуйте перейти по ней — вы попадете на оригинальную страницу.
Другие сервисы для сокращения ссылок
Есть большое количество аналогичных сервисов. Вы можете легко найти их через поиск.
Давайте для примера, я покажу вам такой инструмент, предоставляемый гуглом. Вот он:
Вот он:
Https://goo.gl/
Откроется окно — в нем будет представлена сокращенная ссылка. Скопируйте ее и используйте.
Видео урок: как сделать сокращенную ссылку в ВК
Заключение
Если вы хотите зарабатывать на своей группе вконтакте (см. ), то вы наверняка будете использовать сокращенные ссылки. С их помощью маскируют урл партнерских программ, которые рекламируют владельцы публичных страниц.
Но это не единственный вариант. Любой длинный урл, можно переделать в короткий аналог.
Вопросы?
Вконтакте
В интернете очень часто бывает ситуация, когда необходимо отправить ссылку на тот или иной сайт. И эти ссылки не всегда удобные, а иногда даже выглядят отталкивающе. По этой причине было создано множество сервисов для сокращения ссылок. Сейчас мы рассмотрим самые распространённые сервисы.
Google URL shortener
Простой, но эффективный – всё, что можно сказать об укоротителе от Гугл. Любая длинная ссылка мигом станет коротким адресом.
Для работы с ним переходим на сайт goo.gl и вставляем длинную ссылку в вот такое окно:
Результат отобразится сразу, а чтобы его не потерять, он сохраняется в окошке ниже навсегда.
В это же время стоит отметить один минус – сервис доступен только для старых пользователей начиная с 13 апреля 2018 г.
А с 30 марта 2019 Google полностью закроет этот сервис. По словам разработчиков, они собираются разработать нечто более удобное и в разы лучше теперешнего укоротителя.
Сокращение ссылок ВК
Мало кому известно, но популярная соц. сеть также имеет такую функцию. Кроме обычного укорачивания линка, ВК предоставляет возможность посмотреть на статистику переходов. Её можно сделать как общедоступной, так и личной.
Чтобы сделать ссылку короткой на ВК, переходим сюда vk.com/cc и вставляем URL в окошко.
На примере, мы сократили линк «https://vk.com/cc », и получили доступ к общей статистике переходов по ней.
Кликер
Аналогичный сайт с таким же функционалом. Кликер отличается тем , что после обрезания линка даёт ещё и QR-код. Это удобно, если их некуда сохранить или записать. Код можно сохранить или распечатать, и в миг переходить на нужный ресурс.
Кликер отличается тем , что после обрезания линка даёт ещё и QR-код. Это удобно, если их некуда сохранить или записать. Код можно сохранить или распечатать, и в миг переходить на нужный ресурс.
To.click
Те же функции, только в слегка другом дизайне. В два клика на to.click можно получить короткий адрес.
Сокращатель Алиэкспресс
Наверное, каждый слышал об этом китайском интернет-магазине. И каждый сталкивался с длинными адресами, которые занимают больше места чем текст сообщения.
При этом, не все в курсе о том, что у Aliexpress есть свой сокращатель — ali.onl .
Очень удобно для того, чтобы делиться товарами с друзьями.
Utka.Su
«Утка» позволяет сохранить линки не только текстом, но и QR-кодами.
Также даёт возможность отслеживать все данные о переходах. Пожалуй, один из самых молодых проектов в списке.
Все сайты для обрезки ссылок практически идентичны. Кроме основной функции бывают и дополнения в виде отслеживания перехода, и даже заработка на переходах. Но факт есть факт – отличия минимальны.
Кроме основной функции бывают и дополнения в виде отслеживания перехода, и даже заработка на переходах. Но факт есть факт – отличия минимальны.
Также он может быть полезен при попытке передать ссылку, которую социальная сеть VK.COM считает вредоносной:
Ссылка на подозрительный сайт
Ссылка, по которой Вы попытались перейти, может вести на сайт, который был создан с целью обмана пользователей ВКонтакте и получения за счет этого прибыли.
ВКонтакте всегда заботится о Вашей безопасности!
Часто это предупреждение не соответствует действительности и ВК просто блокирует белые сайты, для обхода этого ограничения и существуют сервисы сокращения ссылок.
Для перехода в сокращатель ссылок от вконтакте вам нужно перейти по одной из этих ссылок –
https://vk.com/cc
https://vk.сс/
На этой странице Вы можете сделать из длинной и сложной ссылки простую.
Такие ссылки удобнее использовать в Ваших записях и сообщениях.
Затем нажмите “получить короткий вариант ссылки”.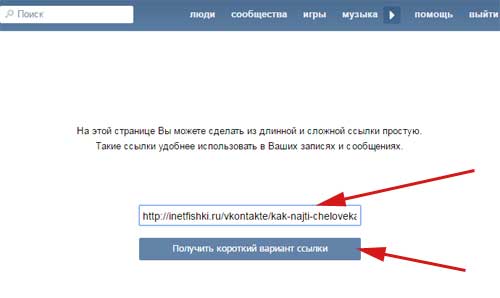
Для чего можно и нужно использовать сокращение ссылок через vk.com/cc:
Не очень красивая, не правда ли? А вот как она выглядит в сокращенном виде – http://vk.cc/3HkKrK
2. Такие ссылки используются для скрытия реферальных ссылок. Чтобы не было надписи “r/18182”, “ref47474” и аналогичных реф. ссылку пропускают через сервис сокращения URL и она становится короткой и не вызывает подозрений
3. Для спама. Часто через многочисленные сервисы сокращения урлов спамеры маскируют ссылки. Сейчас вконтакте уже без проблем расшифровывает “шорт урлы” и банит тех, кто распространяет эти ссылки, даже в сокращенном виде.
5. Для увеличения места в “твите” в твиттере, так как там есть ограничение на число символов в одном посте и вставив длинную ссылку у вас может просто не остаться букв для ее описания.
Множество других способов, когда вам потребуется урезать ссылку.
На самом деле сайтов, предлагающих сократить вам ссылку очень много. Я дам вам адреса нескольких которыми пользуюсь сам:
https://bitly. com/
com/
Если остались вопросы – спрашивайте.
Сервис Vk.cc — сокращение ссылки «ВКонтакте»
Социальная сеть «ВКонтакте» в наши дни используется не только для общения. Многие активно зарабатывают на популярном ресурсе, используя его многочисленные функции. В этой статье вы узнаете, что такое сокращение ссылки ВКонтакте и для чего оно нужно.
Почему мне нужно сокращать ссылки?
Допустим, вам нужно продвинуть свой сайт в социальной сети. Однако большинство ссылок на сторонние ресурсы имеют очень длинные названия, которые в тексте объявления не выглядят привлекательными для потенциального клиента.Уменьшение ссылки «ВКонтакте» придаст ей более компактный вид. К тому же у большинства пользователей даже нет подозрений, что вы имеете в виду сторонний ресурс, а не страницу в социальной сети.
Это активно используется рефералами, которые набирают большую команду рефералов для своих проектов. Однако переходы на сторонние сайты могут привести к мошенническим ресурсам. Поэтому убедитесь, что на вашем компьютере установлена хорошая антивирусная программа. Ведь сокращение ссылки «ВКонтакте» не защитит вас от вредоносных сайтов.Визуально даже профессионал не может определить, куда именно приведет оставленный в сети короткий адрес.
Поэтому убедитесь, что на вашем компьютере установлена хорошая антивирусная программа. Ведь сокращение ссылки «ВКонтакте» не защитит вас от вредоносных сайтов.Визуально даже профессионал не может определить, куда именно приведет оставленный в сети короткий адрес.
Как сократить ссылку?
Чтобы сделать длинную ссылку короткой, вы можете использовать одну из служб в сети. На сегодняшний день эта услуга предоставляется совершенно бесплатно и доступна абсолютно каждому. Просто скопируйте адресную строку в поле, и система выдаст короткую ссылку через несколько секунд.
Уменьшение ссылок «ВКонтакте» (vk.cc) — самый популярный способ среди веб-мастеров, продвигающих собственные сайты или интернет-магазины.Ведь результатом является адрес электронной почты страницы, который очень похож на внутренний переход в социальной сети. Кроме того, такую ссылку можно вставить в якорь и зашифровать под названием «Подробнее» или «Узнать подробнее». Не исключено, что вы встречали такую уловку от создателей групп и паблика в социальной сети.
Сервис сокращения ссылок «ВКонтакте»
Эта функция в популярной социальной сети появилась совсем недавно, но уже успела принести плоды вебмастерам.Чтобы воспользоваться услугой сокращения ссылок, перейдите по адресу vk.cc. Страница представляет собой всего лишь адресную строку, после заполнения которой пользователь получит сокращенный вариант своей ссылки. Вы можете использовать этот результат не только в социальной сети, но и на любом сайте, требующем шифрования. Система не хранит введенные данные, что гарантирует полную конфиденциальность пользователя.
Таким образом, аббревиатура ссылки «ВКонтакте» — это отличный способ быстро и надежно продвинуть свой проект и привлечь как можно больше потенциальных клиентов.В конце концов, будущее за информационными технологиями. А это значит, что нужно использовать все возможности, которые дают нам социальные сети.
p>% PDF-1.7
%
7440 0 объект
>
эндобдж
xref
7440 89
0000000016 00000 н.
0000007062 00000 н. 0000007465 00000 н.
0000007502 00000 н.
0000007588 00000 н.
0000007665 00000 н.
0000007698 00000 п.
0000007799 00000 н.
0000007828 00000 н.
0000008064 00000 н.
0000008853 00000 н.
0000009002 00000 н.
0000009041 00000 н.
0000009090 00000 н.
0000009139 00000 п.
0000009288 00000 п.
0000009401 00000 п.
0000009516 00000 н.
0000009615 00000 н.
0000009741 00000 н.
0000009856 00000 н.
0000009981 00000 н.
0000010108 00000 п.
0000010236 00000 п.
0000010335 00000 п.
0000012510 00000 п.
0000014491 00000 п.
0000016499 00000 н.
0000018571 00000 п.
0000020792 00000 п.
0000022949 00000 п.
0000023325 00000 п.
0000023673 00000 п.
0000025701 00000 п.
0000025945 00000 п.
0000026172 00000 п.
0000027973 00000 п.
0000028097 00000 п.
0000028246 00000 п.
0000028370 00000 п.
0000028519 00000 п.
0000028907 00000 п.
0000029138 00000 п.
0000029252 00000 п.
0000029640 00000 п.
0000032135 00000 п.
0000032191 00000 п.
0000032579 00000 п.
0000032632 00000 п.
0000035282 00000 п.
0000007465 00000 н.
0000007502 00000 н.
0000007588 00000 н.
0000007665 00000 н.
0000007698 00000 п.
0000007799 00000 н.
0000007828 00000 н.
0000008064 00000 н.
0000008853 00000 н.
0000009002 00000 н.
0000009041 00000 н.
0000009090 00000 н.
0000009139 00000 п.
0000009288 00000 п.
0000009401 00000 п.
0000009516 00000 н.
0000009615 00000 н.
0000009741 00000 н.
0000009856 00000 н.
0000009981 00000 н.
0000010108 00000 п.
0000010236 00000 п.
0000010335 00000 п.
0000012510 00000 п.
0000014491 00000 п.
0000016499 00000 н.
0000018571 00000 п.
0000020792 00000 п.
0000022949 00000 п.
0000023325 00000 п.
0000023673 00000 п.
0000025701 00000 п.
0000025945 00000 п.
0000026172 00000 п.
0000027973 00000 п.
0000028097 00000 п.
0000028246 00000 п.
0000028370 00000 п.
0000028519 00000 п.
0000028907 00000 п.
0000029138 00000 п.
0000029252 00000 п.
0000029640 00000 п.
0000032135 00000 п.
0000032191 00000 п.
0000032579 00000 п.
0000032632 00000 п.
0000035282 00000 п. 0000035670 00000 п.
0000035789 00000 п.
0000035938 00000 п.
0000046137 00000 п.
0000046178 00000 п.
0000046255 00000 п.
0000046736 00000 п.
0000046813 00000 п.
0000047257 00000 п.
0000047334 00000 п.
0000047821 00000 п.
0000047898 00000 н.
0000047930 00000 н.
0000048007 00000 п.
0000049808 00000 п.
0000050136 00000 п.
0000050205 00000 п.
0000050323 00000 п.
0000052124 00000 п.
0000055622 00000 п.
0000056001 00000 п.
0000056078 00000 п.
0000056492 00000 п.
0000056569 00000 п.
0000056834 00000 п.
0000056884 00000 п.
0000056916 00000 п.
0000056993 00000 п.
0000059087 00000 п.
0000059410 00000 п.
0000059479 00000 п.
0000059597 00000 п.
0000061691 00000 п.
0000072176 00000 п.
0000072534 00000 п.
0000072611 00000 п.
0000342199 00000 п.
0000006765 00000 н.
0000002117 00000 н.
трейлер
] / Назад 1538549 / XRefStm 6765 >>
startxref
0
%% EOF
7528 0 объект
> поток
hYy \ S? & 4! @ «* Pi @ Vp4jDq \ m / An @ +} * Ԫ`
Q [VgʢU [soN ٿ {
0000035670 00000 п.
0000035789 00000 п.
0000035938 00000 п.
0000046137 00000 п.
0000046178 00000 п.
0000046255 00000 п.
0000046736 00000 п.
0000046813 00000 п.
0000047257 00000 п.
0000047334 00000 п.
0000047821 00000 п.
0000047898 00000 н.
0000047930 00000 н.
0000048007 00000 п.
0000049808 00000 п.
0000050136 00000 п.
0000050205 00000 п.
0000050323 00000 п.
0000052124 00000 п.
0000055622 00000 п.
0000056001 00000 п.
0000056078 00000 п.
0000056492 00000 п.
0000056569 00000 п.
0000056834 00000 п.
0000056884 00000 п.
0000056916 00000 п.
0000056993 00000 п.
0000059087 00000 п.
0000059410 00000 п.
0000059479 00000 п.
0000059597 00000 п.
0000061691 00000 п.
0000072176 00000 п.
0000072534 00000 п.
0000072611 00000 п.
0000342199 00000 п.
0000006765 00000 н.
0000002117 00000 н.
трейлер
] / Назад 1538549 / XRefStm 6765 >>
startxref
0
%% EOF
7528 0 объект
> поток
hYy \ S? & 4! @ «* Pi @ Vp4jDq \ m / An @ +} * Ԫ`
Q [VgʢU [soN ٿ {
Новые данные о старении и связанных со старением заболеваниях у эукариотических и прокариотических модельных организмов
Старение связано с накоплением клеточных повреждений в течение жизни. Этому процессу в значительной степени способствуют активные формы кислорода (АФК), образующиеся через клеточные метаболические и дыхательные пути. Фармакологические, нефармакологические и генетические вмешательства использовались для нацеливания на клеточные и митохондриальные сети в попытке расшифровать старение и связанные с возрастом расстройства. В то время как ROS исторически рассматривались как вредный побочный продукт нормального метаболизма и связаны с несколькими патологиями, недавние исследования показали более сложную и полезную роль ROS в регулировании метаболизма, развития и продолжительности жизни.В этом обзоре мы суммируем последние достижения в исследованиях АФК, уделяя особое внимание положительной и вредной роли АФК, многие из которых сохраняются у разных видов, от бактерий до людей, в различных аспектах клеточной физиологии. Эти исследования предоставляют новый контекст для нашего понимания роли АФК в здоровье и болезнях. Кроме того, мы подчеркиваем полезность бактериальных моделей для выяснения молекулярных путей, с помощью которых АФК опосредуют старение и связанные со старением заболевания.
Этому процессу в значительной степени способствуют активные формы кислорода (АФК), образующиеся через клеточные метаболические и дыхательные пути. Фармакологические, нефармакологические и генетические вмешательства использовались для нацеливания на клеточные и митохондриальные сети в попытке расшифровать старение и связанные с возрастом расстройства. В то время как ROS исторически рассматривались как вредный побочный продукт нормального метаболизма и связаны с несколькими патологиями, недавние исследования показали более сложную и полезную роль ROS в регулировании метаболизма, развития и продолжительности жизни.В этом обзоре мы суммируем последние достижения в исследованиях АФК, уделяя особое внимание положительной и вредной роли АФК, многие из которых сохраняются у разных видов, от бактерий до людей, в различных аспектах клеточной физиологии. Эти исследования предоставляют новый контекст для нашего понимания роли АФК в здоровье и болезнях. Кроме того, мы подчеркиваем полезность бактериальных моделей для выяснения молекулярных путей, с помощью которых АФК опосредуют старение и связанные со старением заболевания.
1. Введение
Старение характеризуется постепенной потерей физической формы с течением времени.Старение проявляется как серия динамических изменений на молекулярном и макромолекулярном уровне в течение жизни [1]. Неправильная регуляция клеточных процессов может повредить физиологическую целостность клетки и впоследствии привести к накоплению поврежденных побочных продуктов. Человечество было очаровано получением лучшего понимания старения на протяжении многих веков, но точные механизмы, лежащие в основе процесса старения человека, остаются в значительной степени неясными. Сам процесс старения сложен из-за нескольких факторов, таких как факторы окружающей среды, социально-экономический статус, физические характеристики и образ жизни [2].
За последние несколько десятилетий ожидаемая продолжительность жизни во всем мире линейно увеличивалась до в среднем 60 лет. Ожидается, что население мира старше 60 лет увеличится с примерно 900 миллионов человек (12%) в 2015 году до примерно 2 миллиардов человек (22%) в 2050 году [3]. Такое увеличение продолжительности жизни связано, в частности, со снижением детской смертности, повышением уровня жизни и медицинскими достижениями. Несмотря на увеличение общей продолжительности жизни, старение и возрастные заболевания являются основными причинами смертности и заболеваемости во всем мире [4].Более того, возрастные расстройства, такие как болезнь Альцгеймера, деменция, сердечно-легочные расстройства, диабет, нейродегенеративные и когнитивные нарушения, хрупкое физическое состояние и психосоматические расстройства, являются основными причинами инвалидности во всем мире. На эти нарушения приходится более 20% лет, прожитых с инвалидностью [5]. Понимание молекулярных механизмов старения имеет решающее значение для разработки терапевтических вмешательств, способствующих здоровому старению.
Такое увеличение продолжительности жизни связано, в частности, со снижением детской смертности, повышением уровня жизни и медицинскими достижениями. Несмотря на увеличение общей продолжительности жизни, старение и возрастные заболевания являются основными причинами смертности и заболеваемости во всем мире [4].Более того, возрастные расстройства, такие как болезнь Альцгеймера, деменция, сердечно-легочные расстройства, диабет, нейродегенеративные и когнитивные нарушения, хрупкое физическое состояние и психосоматические расстройства, являются основными причинами инвалидности во всем мире. На эти нарушения приходится более 20% лет, прожитых с инвалидностью [5]. Понимание молекулярных механизмов старения имеет решающее значение для разработки терапевтических вмешательств, способствующих здоровому старению.
Митохондрии, которые часто называют «электростанцией клетки», метаболизируют углеводы и жирные кислоты посредством окислительного фосфорилирования.Благодаря этому процессу митохондрии могут генерировать от 32 до 34 молекул аденозинтрифосфата (АТФ) на молекулу глюкозы. Белковые комплексы во внутренней митохондриальной мембране вместе образуют митохондриальную цепь переноса электронов (ETC), которая высвобождает свободные радикалы в качестве побочных продуктов энергетического метаболизма [6]. Харман первоначально предложил свободнорадикальную теорию старения в 1956 году [7], согласно которой активные формы кислорода (АФК) являются основными медиаторами процесса старения. Краткий обзор источников АФК и последующих клеточных ответов представлен на рисунке 1.Источники АФК, антиоксидантной защиты и последующих биологических эффектов были рассмотрены в другом месте (например, [8]) и не будут подробно рассматриваться в этом обзоре. В то время как обширные данные указывают на то, что повышенная продукция ROS и сниженная способность поглощать ROS сокращают продолжительность жизни [9, 10], теория старения со свободными радикалами столкнулась с противодействием, подрывая идею о том, что только ROS несут ответственность за процесс старения. Например, организмы могут жить здоровой жизнью в отсутствие поглотителей АФК [11–14].
Белковые комплексы во внутренней митохондриальной мембране вместе образуют митохондриальную цепь переноса электронов (ETC), которая высвобождает свободные радикалы в качестве побочных продуктов энергетического метаболизма [6]. Харман первоначально предложил свободнорадикальную теорию старения в 1956 году [7], согласно которой активные формы кислорода (АФК) являются основными медиаторами процесса старения. Краткий обзор источников АФК и последующих клеточных ответов представлен на рисунке 1.Источники АФК, антиоксидантной защиты и последующих биологических эффектов были рассмотрены в другом месте (например, [8]) и не будут подробно рассматриваться в этом обзоре. В то время как обширные данные указывают на то, что повышенная продукция ROS и сниженная способность поглощать ROS сокращают продолжительность жизни [9, 10], теория старения со свободными радикалами столкнулась с противодействием, подрывая идею о том, что только ROS несут ответственность за процесс старения. Например, организмы могут жить здоровой жизнью в отсутствие поглотителей АФК [11–14]. Кроме того, пищевые, фармакологические и генетические вмешательства, которые увеличивают производство АФК, могут способствовать долголетию за счет активации митохондриального окислительного фосфорилирования и запуска нижестоящих сигнальных путей, которые способствуют адаптивному ответу [11, 15, 16], в то время как фармакологические вмешательства, ограничивающие производство АФК, были показаны. сократить продолжительность жизни [11, 15].
Кроме того, пищевые, фармакологические и генетические вмешательства, которые увеличивают производство АФК, могут способствовать долголетию за счет активации митохондриального окислительного фосфорилирования и запуска нижестоящих сигнальных путей, которые способствуют адаптивному ответу [11, 15, 16], в то время как фармакологические вмешательства, ограничивающие производство АФК, были показаны. сократить продолжительность жизни [11, 15].
Хотя окислительное повреждение, вызванное ROS и ROS, не может быть единственной причиной процесса старения, вполне очевидно, что ROS действительно играют важную роль в молекулярных механизмах, влияющих на продолжительность жизни.Таким образом, устранение разрыва между теорией свободных радикалов и современными знаниями о старении может помочь нам лучше понять, как взаимодействие между окислительным повреждением, вызванным АФК, и клеточным метаболизмом влияет на старение, и раскрыть генетические и фармацевтические меры, которые могут модулировать это взаимодействие.
2. Свободнорадикальная теория старения и за ее пределами
За последние несколько десятилетий доминирующей моделью старения была свободнорадикальная теория старения. Эта теория утверждает, что организмы стареют, потому что они накапливают окислительные повреждения, вызванные ROS.АФК представляют собой частично восстановленные метаболиты молекулярного кислорода, генерируемые различными метаболическими реакциями и клеточными процессами, такими как дыхание [11, 15]. Несколько исследований подтверждают теорию старения, основанную на свободных радикалах. Например, было показано, что производные чеснока тиоаллильные соединения S-аллилцистеин и S-аллилмеркаптоцистеин уменьшают накопление ROS и увеличивают продолжительность жизни C. elegans [17]. Точно так же обработка C. elegans четырьмя синтетическими производными стильбена увеличивала продолжительность жизни за счет снижения накопления ROS и окислительного стресса [18].
Однако недавние исследования показывают, что АФК играют более сложную роль в определении долголетия, чем считалось ранее. Например, мутанты C. elegans , лишенные супероксиддисмутазы (SOD) — фермента, нейтрализующего супероксидный радикал, — будучи более восприимчивыми к множественным стрессорам, сохраняют нормальную продолжительность жизни [12]. В другом исследовании было обнаружено, что делеция митохондриальной супероксиддисмутазы sod-2 действительно увеличивает продолжительность жизни C. elegans [19].Кроме того, у C. elegans отсутствуют функциональные гены субъединиц комплексов I и III митохондриальной дыхательной цепи, продуцирующие более высокие уровни супероксида, но они также имеют увеличенную продолжительность жизни. Увеличение продолжительности жизни этих нокаутов может быть полностью отменено обработкой их поглотителем супероксида N -ацетилцистеином [20]. Кроме того, когда дикий тип C. elegans и долгоживущий митохондриальный мутант clk-1 обрабатывали паракватом, генератором супероксида, как средняя, так и максимальная продолжительность жизни значительно увеличивались [20].
Например, мутанты C. elegans , лишенные супероксиддисмутазы (SOD) — фермента, нейтрализующего супероксидный радикал, — будучи более восприимчивыми к множественным стрессорам, сохраняют нормальную продолжительность жизни [12]. В другом исследовании было обнаружено, что делеция митохондриальной супероксиддисмутазы sod-2 действительно увеличивает продолжительность жизни C. elegans [19].Кроме того, у C. elegans отсутствуют функциональные гены субъединиц комплексов I и III митохондриальной дыхательной цепи, продуцирующие более высокие уровни супероксида, но они также имеют увеличенную продолжительность жизни. Увеличение продолжительности жизни этих нокаутов может быть полностью отменено обработкой их поглотителем супероксида N -ацетилцистеином [20]. Кроме того, когда дикий тип C. elegans и долгоживущий митохондриальный мутант clk-1 обрабатывали паракватом, генератором супероксида, как средняя, так и максимальная продолжительность жизни значительно увеличивались [20].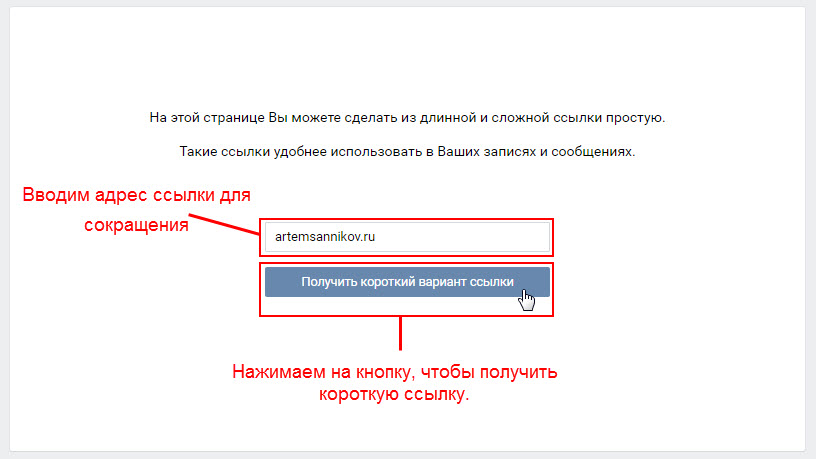
2.1. Антиоксидантные ферменты: хорошо или плохо?
Антиоксидантные ферменты играют ключевую роль в нейтрализации различных АФК. Однако взаимосвязь между уровнями антиоксидантных ферментов и продолжительностью жизни не является однозначной.
Несколько исследований, посвященных роли системы антиоксидантной защиты в регулировании долголетия, показали, что повышенная устойчивость к окислительному стрессу может улучшить продолжительность жизни мышей [21, 22]. Например, мыши с нокаутом Cu / Zn супероксиддисмутазы 1 ( Sod1 — / — ) значительно сократили продолжительность жизни.Это сокращение продолжительности жизни было связано с повышенным клеточным старением на основе повышенной экспрессии маркеров старения p16 и p21 [23]. Кроме того, избыточная экспрессия митохондриальной каталазы связана с увеличением средней и максимальной продолжительности жизни у трансгенных мышей, избыточно экспрессирующих пероксисомальную, ядерную и митохондриальную каталазы [24]. Также было показано, что сверхэкспрессия митохондриальной каталазы снижает различные возрастные патологические состояния, такие как сердечные проблемы, заболевания, связанные с воспалением, и рак [25] .
Также было показано, что сверхэкспрессия митохондриальной каталазы снижает различные возрастные патологические состояния, такие как сердечные проблемы, заболевания, связанные с воспалением, и рак [25] .
Однако другие исследования показали, что повышенная активность антиоксидантных ферментов не способствует увеличению продолжительности жизни у грызунов [26]. Например, исследование трансгенных мышей, сверхэкспрессирующих Cu / Zn SOD, Mn-SOD и каталазу, по отдельности или в комбинации, показало, что сверхэкспрессия этих ферментов не приводит к значительному увеличению продолжительности жизни по сравнению с мышами дикого типа (WT) [27]. .
Глутатионпероксидаза 1 (GPX1), основная изоформа семейства белков GPX, является важным антиоксидантным ферментом, который присутствует в клетках повсеместно и играет важную роль в нейтрализации перекиси водорода.В то время как экспрессия GPX1 имеет защитный эффект против ROS-опосредованного клеточного повреждения, мыши Gpx1 с нокаутом не показали никаких доказательств повышенного окислительного повреждения белков и липидов по сравнению с их однопометниками WT [28]. Напротив, мыши, лишенные как Mn-SOD, так и Gpx1 , имели более высокий уровень окислительного повреждения ДНК и белков, но их продолжительность жизни не сокращалась по сравнению с однопометниками дикого типа [29]. Более того, однонуклеотидный полиморфизм генов Mn-SOD и Gpx1 , как было показано, влияет на старение и долголетие [30].
Напротив, мыши, лишенные как Mn-SOD, так и Gpx1 , имели более высокий уровень окислительного повреждения ДНК и белков, но их продолжительность жизни не сокращалась по сравнению с однопометниками дикого типа [29]. Более того, однонуклеотидный полиморфизм генов Mn-SOD и Gpx1 , как было показано, влияет на старение и долголетие [30].
Другой белок семейства GPX, GPX4, играет важную роль в защите плазматической мембраны от вызванного пероксидом повреждения липидов. Нулевые мутации гена Gpx4 смертельны для мышей. Удаление GPX4 у линии трансгенных мышей (фон C57BL / 6) приводило к усилению окислительного повреждения мозга, а также к потере нейронов по сравнению с мышами WT [31]. Было показано, что трансгенная сверхэкспрессия GPX4 защищает мышей от фенотипа летальных нуль-мутаций и предотвращает вызванное окислительным стрессом повреждение печени и гибель клеток [32].Однако мыши со сниженной экспрессией и активностью GPX4 не показали значительных различий в средней, средней и максимальной продолжительности жизни по сравнению с мышами WT [33].
Тиоредоксин (Trx) — это редокс-белок, который действует как донор водорода во многих восстановительных реакциях в клетках. Он имеет две формы: цитоплазматическую (Trx1) и митохондриальную (Trx2). Подобно Gpx4 , нулевые мутации Trx2 являются летальными для мышей [34], а нокаут Trx2 нарушает функцию митохондрий за счет снижения продукции АТФ; увеличение производства АФК; индуцирование окислительного повреждения ДНК, белков и липидов в печени; и увеличение апоптоза клеток печени, вызванного окислительным стрессом [35].Было показано, что сверхэкспрессия Trx1 (Tg ( TRX1 ) + / 0 ) защищает от окислительного повреждения клеточных макромолекул и продлевает более раннюю часть продолжительности жизни у самцов мышей; однако ни самцы, ни самки мышей Tg ( TRX1 ) + / 0 не показали изменений максимальной продолжительности жизни [36].
Местоположение сотовой связи, где производится производство ROS, может определять, играют ли ROS положительную или отрицательную роль. Например, делеция митохондрий sod-2 в C.elegans , как было показано, способствует долголетию, тогда как делеция цитоплазматических sod-1 и sod-5 ограничивает продолжительность жизни [37]. Было показано, что АФК, продуцируемые обратным транспортом электронов митохондриального респираторного комплекса I, увеличивают продолжительность жизни у Drosophila [38]. Более того, ингибирование дыхания, по-видимому, активирует индуцируемый гипоксией фактор-1 (HIF-1) за счет повышения уровней ROS. Было показано, что эта активация увеличивает продолжительность жизни [17, 18]. Исследования на генетически модифицированных мышах показали, что умеренно нарушенная функция митохондрий может привести к более здоровому старению, тогда как сильно измененный митохондриальный гомеостаз может быть пагубным [39, 40].На основании этих наблюдений становится ясно, что как уровень, так и расположение продукции ROS вносят вклад в определение роли ROS в регуляции долголетия [41].
Например, делеция митохондрий sod-2 в C.elegans , как было показано, способствует долголетию, тогда как делеция цитоплазматических sod-1 и sod-5 ограничивает продолжительность жизни [37]. Было показано, что АФК, продуцируемые обратным транспортом электронов митохондриального респираторного комплекса I, увеличивают продолжительность жизни у Drosophila [38]. Более того, ингибирование дыхания, по-видимому, активирует индуцируемый гипоксией фактор-1 (HIF-1) за счет повышения уровней ROS. Было показано, что эта активация увеличивает продолжительность жизни [17, 18]. Исследования на генетически модифицированных мышах показали, что умеренно нарушенная функция митохондрий может привести к более здоровому старению, тогда как сильно измененный митохондриальный гомеостаз может быть пагубным [39, 40].На основании этих наблюдений становится ясно, что как уровень, так и расположение продукции ROS вносят вклад в определение роли ROS в регуляции долголетия [41].
3. Роль АФК в повреждении ядерной и митохондриальной ДНК
Повреждение ядерной и митохондриальной ДНК, вызванное АФК, вносит значительный вклад в процесс старения. В нормальных физиологических условиях множество механизмов восстановления ДНК работают в гармонии, чтобы сдерживать повреждение. Базовая эксцизионная репарация, репарация ошибочного спаривания, эксцизионная репарация нуклеотидов и репарация двухцепочечного разрыва — все это работает строго для исправления повреждений ДНК, вызванных ROS, рентгеновскими лучами, ультрафиолетовым и ионизирующим излучением, щелочными агентами, ошибками репликации, противоопухолевыми агентами и различными химическими веществами. агенты [42].Недостаток любого из этих механизмов восстановления может ускорить начало старения [43].
ДНК-теория старения, впервые предложенная Сциллардом в 1959 году [44], коррелирует устойчивое накопление повреждений ДНК с дисбалансом клеточной функции, что в конечном итоге приводит к старению клеток и организма. Виленчик и Кнудсон [45] подсчитали, что геном млекопитающих может выдерживать до 1000 повреждений в час на клетку. Эти поражения включают окислительное повреждение оснований, поперечных связей и одно- / двухцепочечные разрывы.Эндогенные АФК обычно вызывают образование абазических сайтов за счет разрыва гликозидных связей между нуклеотидными основаниями и остатками дезоксирибозы [46, 47]. Агенты окружающей среды, такие как УФ-лучи и химические мутагены, вызывают разрывы цепей из-за модификаций оснований и интеркаляций [48, 49]. Когда не восстановленные повреждения накапливаются, это запускает реакцию на повреждение ДНК (DDR) [50, 51], которая активирует системы репарации ДНК [43]. Несмотря на количество поражений, от которых страдает геном, частота реальных мутаций намного ниже именно из-за этих хорошо скоординированных систем восприятия и восстановления.Однако, когда механизмы репарации ДНК нарушены или становятся дисфункциональными, DDR запускает старение или апоптоз, чтобы приостановить или устранить поврежденные клетки, соответственно.
Виленчик и Кнудсон [45] подсчитали, что геном млекопитающих может выдерживать до 1000 повреждений в час на клетку. Эти поражения включают окислительное повреждение оснований, поперечных связей и одно- / двухцепочечные разрывы.Эндогенные АФК обычно вызывают образование абазических сайтов за счет разрыва гликозидных связей между нуклеотидными основаниями и остатками дезоксирибозы [46, 47]. Агенты окружающей среды, такие как УФ-лучи и химические мутагены, вызывают разрывы цепей из-за модификаций оснований и интеркаляций [48, 49]. Когда не восстановленные повреждения накапливаются, это запускает реакцию на повреждение ДНК (DDR) [50, 51], которая активирует системы репарации ДНК [43]. Несмотря на количество поражений, от которых страдает геном, частота реальных мутаций намного ниже именно из-за этих хорошо скоординированных систем восприятия и восстановления.Однако, когда механизмы репарации ДНК нарушены или становятся дисфункциональными, DDR запускает старение или апоптоз, чтобы приостановить или устранить поврежденные клетки, соответственно.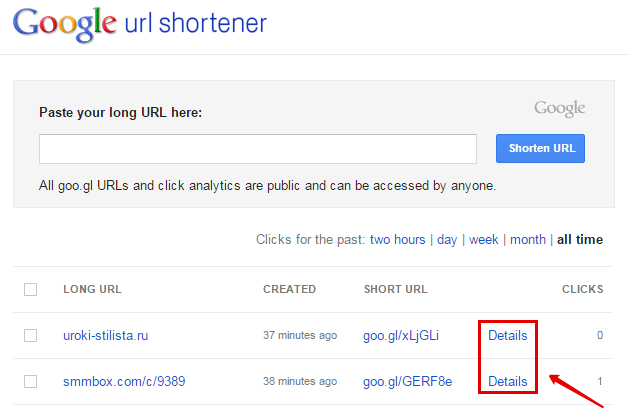 Накопление стареющих клеток в стареющих тканях [32] считается движущей силой процесса старения, в первую очередь через воспалительные пути [33].
Накопление стареющих клеток в стареющих тканях [32] считается движущей силой процесса старения, в первую очередь через воспалительные пути [33].
Реставрацию ДНК можно разделить на три типа: эксцизионная репарация оснований (BER), эксцизионная репарация нуклеотидов (NER) и негомологичное соединение концов (NHEJ). Эти процессы подробно рассмотрены в литературе [52–54].BER обычно восстанавливает окислительное повреждение ДНК, чаще всего поражение 8-оксогуанином [55]. Вкратце, ДНК-гликозилазы вырезают поврежденное основание, а полимераза вставляет правильный нуклеотид на его место [56]. NER корректирует более сложные поражения, не связанные с окислительным повреждением, такие как образование аддукта между основаниями и индуцированные ультрафиолетовыми лучами перекрестные связи [57]. В то время как эксцизионная репарация в основном происходит во время репликации, NHEJ может восстанавливать двухцепочечные разрывы ДНК также и в состоянии покоя [58]. NHEJ — это трехэтапный процесс, который начинается со связывания конца разорванной цепи с белком Ku. Поврежденные и / или несовпадающие нуклеотиды затем удаляются, и правильная последовательность синтезируется ДНК-полимеразой [59].
Поврежденные и / или несовпадающие нуклеотиды затем удаляются, и правильная последовательность синтезируется ДНК-полимеразой [59].
Неудивительно, что исследования наблюдали связанное с возрастом снижение уровней и активности белка репарации ДНК [55]. Сообщалось о снижении активности BER в различных тканях у пожилых людей [60] и у мышей, лишенных сиртуина 6, гистондеацетилазы, которая активна во время репарации ДНК [61]. Снижение уровня белка Ku и других медиаторов NHEJ наблюдается при нормальном старении человека и в случаях болезни Альцгеймера [62].Сходным образом активность NHEJ также снижается у старых крыс, у которых накопились разрывы цепей ДНК в своих нейронах [63].
Наиболее убедительные доказательства ДНК-теории старения дают человеческие прогероидные синдромы (то есть преждевременное старение), такие как синдром Вернера (WS), синдром Блума (BS) и пигментная ксеродермия (XP). Эти синдромы вызваны нестабильностью генома и основным дефектом репарации ДНК. WS и BS вызваны мутациями потери функции в генах WRN и BLM соответственно [64, 65]. Эти гены кодируют геликазы RecQ, которые участвуют как в репликации, так и в репарации ДНК и, как известно, взаимодействуют с белком Ku [66, 67]. Нокауты WRN и BLM у мышей обладают значительной геномной нестабильностью и нарушенными механизмами репарации ДНК по сравнению с мышами WT [68, 69]. XP характеризуется мутацией в нуклеазе эксцизионной репарации кросс-комплементации группы 1 xeroderma pigmentosum group F (ERCC1-XPF), которая играет важную роль в репарации как NER, так и NHEJ [70]. У мышей, лишенных ERCC1 , наблюдается ускоренное старение кожи, повышенное повреждение ДНК и клеточное старение по сравнению с мышами WT [71].Репликативное укорочение теломер было вовлечено в старение на основании исследований на мышиной модели с нокаутом теломеразы. Эта модель мышей демонстрирует прогерию и накапливает обширные повреждения ДНК (обзор [72]). Укорочение теломер также сопровождает синдромы прогерии у человека, такие как WS и BS. Более поздние исследования прямо связали дефектную репарацию ДНК, особенно BER и NER, с сайтами DDR, вызванного распаковкой теломер [73, 74].
Эти гены кодируют геликазы RecQ, которые участвуют как в репликации, так и в репарации ДНК и, как известно, взаимодействуют с белком Ku [66, 67]. Нокауты WRN и BLM у мышей обладают значительной геномной нестабильностью и нарушенными механизмами репарации ДНК по сравнению с мышами WT [68, 69]. XP характеризуется мутацией в нуклеазе эксцизионной репарации кросс-комплементации группы 1 xeroderma pigmentosum group F (ERCC1-XPF), которая играет важную роль в репарации как NER, так и NHEJ [70]. У мышей, лишенных ERCC1 , наблюдается ускоренное старение кожи, повышенное повреждение ДНК и клеточное старение по сравнению с мышами WT [71].Репликативное укорочение теломер было вовлечено в старение на основании исследований на мышиной модели с нокаутом теломеразы. Эта модель мышей демонстрирует прогерию и накапливает обширные повреждения ДНК (обзор [72]). Укорочение теломер также сопровождает синдромы прогерии у человека, такие как WS и BS. Более поздние исследования прямо связали дефектную репарацию ДНК, особенно BER и NER, с сайтами DDR, вызванного распаковкой теломер [73, 74]. Примеры репарации специфических повреждений ДНК и дефектов реакции, которые приводят к генетическим нарушениям у человека, показаны на рисунке 2.Таким образом, есть существенные доказательства связи нарушения репарации ДНК с синдромами старения; однако необходимы дальнейшие исследования, чтобы установить прямую механистическую связь.
Примеры репарации специфических повреждений ДНК и дефектов реакции, которые приводят к генетическим нарушениям у человека, показаны на рисунке 2.Таким образом, есть существенные доказательства связи нарушения репарации ДНК с синдромами старения; однако необходимы дальнейшие исследования, чтобы установить прямую механистическую связь.
Поскольку митохондрии являются основными участками продукции АФК, митохондриальная ДНК (мтДНК) содержит более высокие уровни окислительного повреждения, а скорость ее мутаций значительно выше, чем у ядерной ДНК [75]. В дополнение к их близости к местам генерации АФК, вероятно, что митохондриальные геномы более склонны к окислительному повреждению, потому что гистоны и другие связанные с хроматином белки, присутствующие в ядерных геномах, где они действуют как поглотители кислородных радикалов, отсутствуют в митохондрии.Существование восстановления окислительного повреждения мтДНК, о котором впервые сообщалось в начале 90-х годов, хорошо установлено [76–78]. BER, по-видимому, является единственным процессом эксцизионной репарации, активным в митохондриальных геномах. Все репарационные белки мтДНК кодируются ядерным геномом и импортируются в митохондриальный матрикс. Большинство обнаруженных к настоящему времени белков репарации мтДНК представляют собой изоформы ядерных белков BER, возникающие в результате дифференциального сплайсинга или усечения концевых последовательностей [79, 80]. Митохондриальная ДНК-полимераза γ (Pol γ ) и лигаза мтДНК (Lig III α ), участвующие в репликации мтДНК, по-видимому, также функционируют в митохондриальной BER [79, 80].
BER, по-видимому, является единственным процессом эксцизионной репарации, активным в митохондриальных геномах. Все репарационные белки мтДНК кодируются ядерным геномом и импортируются в митохондриальный матрикс. Большинство обнаруженных к настоящему времени белков репарации мтДНК представляют собой изоформы ядерных белков BER, возникающие в результате дифференциального сплайсинга или усечения концевых последовательностей [79, 80]. Митохондриальная ДНК-полимераза γ (Pol γ ) и лигаза мтДНК (Lig III α ), участвующие в репликации мтДНК, по-видимому, также функционируют в митохондриальной BER [79, 80].
Было обнаружено, что накопление соматических мутаций мтДНК ускоряет нормальное старение [81–84], что приводит к окислительному повреждению, энергетической недостаточности, увеличению продукции ROS и накоплению пептида бета-амилоида (Abeta) [85, 86], a ключевая молекула при болезни Альцгеймера (БА) [83]. Возникает порочный круг, который усиливает повреждение мтДНК, нарушение митохондриального дыхания и окислительный стресс.
4. Роль АФК в гомеостазе белка
Подобно повреждению ДНК, возрастное повреждение белка и накопление поврежденных белковых продуктов способствуют старению.Поэтому очень важно понять, как АФК вносят вклад в дисбаланс в гомеостазе клеточных белков и изменяют процесс старения.
Свободные радикалы могут «атаковать» белки, вызывая окислительное повреждение. Окислительное повреждение может изменить функцию белка. Кроме того, он может образовывать двойные связи углерод-кислород в боковых цепях аргинина, лизина, пролина и треонина, образуя реактивные кетоны или альдегиды, известные как белковые карбонилы [87], которые обычно считаются отражающими общий уровень клеточного окислительного стресса [88]. .Карбонилы белков связаны с образованием аберрантных изоформ белков [89, 90]. В отличие от других окислительных модификаций, таких как образование дисульфидной связи, карбонилирование белка необратимо. Таким образом, единственным средством ограничения повреждений, вызываемых пораженными белками, является их деградация. Чем больше накапливается окислительное повреждение, тем больше вероятность неправильного складывания белков. Умеренно окисленные белки подвергаются деградации протеасомой, сложным комплексом протеаз, разработанным для осуществления селективной, эффективной и процессной деградации короткоживущих, поврежденных, неправильно свернутых или устаревших белков [53].Однако сильно окисленные белки могут сшиваться с другими белками, что предотвращает их деградацию [54]. Как следствие, внутри клетки накапливаются сильно поврежденные белки, что влияет на ее нормальное функционирование. Соответственно, нарушение протеостаза является признаком многих возрастных заболеваний, включая болезнь Альцгеймера и Паркинсона [91, 92].
Чем больше накапливается окислительное повреждение, тем больше вероятность неправильного складывания белков. Умеренно окисленные белки подвергаются деградации протеасомой, сложным комплексом протеаз, разработанным для осуществления селективной, эффективной и процессной деградации короткоживущих, поврежденных, неправильно свернутых или устаревших белков [53].Однако сильно окисленные белки могут сшиваться с другими белками, что предотвращает их деградацию [54]. Как следствие, внутри клетки накапливаются сильно поврежденные белки, что влияет на ее нормальное функционирование. Соответственно, нарушение протеостаза является признаком многих возрастных заболеваний, включая болезнь Альцгеймера и Паркинсона [91, 92].
Многие исследования показали связь между гомеостазом белков, АФК и окислительным стрессом. Например, было показано, что снижение передачи сигналов инсулина / IGF-1 или ингибирование передачи сигналов нижестоящего mTOR улучшает гомеостаз белков, связанных с болезнью Альцгеймера, способствуя долголетию и защищая когнитивные функции на животных моделях [93]. Несколько исследований на C. elegans также показали, что фактор теплового шока 1 (HSF-1) работает с FOXO-подобным фактором транскрипции, daf-16 , для улучшения гомеостаза белков и увеличения продолжительности жизни [94, 95]. Обработка C. elegans амилоидсвязывающим красителем тиофлавином Т снижает агрегацию белков и увеличивает продолжительность жизни за счет передачи сигналов, опосредованной HSF-1 и SKN-1 / Nrf [96]. Другое исследование, сравнивающее роль малых белков теплового шока у Drosophila , идентифицировало два белка — CG14207 и HSP67BC — участвующих в протеостазе, которые слегка улучшали продолжительность жизни при сверхэкспрессии у Drosophila [97].
Несколько исследований на C. elegans также показали, что фактор теплового шока 1 (HSF-1) работает с FOXO-подобным фактором транскрипции, daf-16 , для улучшения гомеостаза белков и увеличения продолжительности жизни [94, 95]. Обработка C. elegans амилоидсвязывающим красителем тиофлавином Т снижает агрегацию белков и увеличивает продолжительность жизни за счет передачи сигналов, опосредованной HSF-1 и SKN-1 / Nrf [96]. Другое исследование, сравнивающее роль малых белков теплового шока у Drosophila , идентифицировало два белка — CG14207 и HSP67BC — участвующих в протеостазе, которые слегка улучшали продолжительность жизни при сверхэкспрессии у Drosophila [97].
Двумя важными протеолитическими путями являются убиквитин-протеасомный путь (UPP) и аутофагия [98]. UPP — это протеолитическая система, ответственная за большую часть деградации внутриклеточного белка. Ключевым аспектом UPP-опосредованного протеолиза является избирательное нацеливание белков для деградации посредством посттрансляционных модификаций, особенно убиквитинирования и сумоилирования [77, 78]. Старение связано с повышенными уровнями убиквитинированного и сумоилированного белка в различных тканях [99–103], возможно, в результате возрастного нарушения функции UPP [104, 105].
Было показано, что пути убиквитинирования играют важную роль в регуляции продолжительности жизни [106, 107]. У Drosophila мутация потери функции в ubiquitin-активирующем ферменте Uba1 значительно сокращает продолжительность жизни и ослабляет моторную функцию [108]. В C. elegans сверхэкспрессия E3 ubiquitin ligase, WWP-1, увеличивала продолжительность жизни посредством передачи сигналов, опосредованной фактором транскрипции forkhead box A (FoxA) [109].
Повышенная экспрессия протеина сборки протеасомы Ump1 также была связана с повышенной жизнеспособностью после воздействия различных факторов окислительного стресса (например,g., менадион, перекись водорода и 4-гидроксиноненаль) в S. cerevisiae [89]. Эта повышенная жизнеспособность была связана с улучшенным сохранением протеасом-опосредованной деградации белка. Интересно, что клетки, экспрессирующие повышенные уровни Ump1, также демонстрируют повышенное сохранение протеасом-опосредованной деградации белка и повышенную жизнеспособность во время старения в стационарной фазе. Взятые вместе, эти данные убедительно подтверждают ключевую роль протеасомы во время окислительного стресса и старения [89].
Аутофагия также важна для поддержания гомеостаза белков, поскольку как клеточная аутофагия, так и митофагия (аутофагия всей митохондрии) влияют на продолжительность жизни [90, 110]. Три белка аутофагии (LC3B, ATG5 и ATG12) играют важную роль в сохранении целостности митохондрий и продолжительности жизни [111]. Было обнаружено, что в эндотелиальных клетках пупочной вены человека целевое повреждение митохондрий запускает каскад событий, включающих кратковременное увеличение продукции ROS, за которым следует митохондриальная фрагментация и активация LC3B, ATG5 и ATG12.Этот каскад значительно увеличил репликативную продолжительность жизни до 150% и количество удвоений популяции до 200% [111]. Кроме того, при нормальном старении и прогрессировании возрастных патологий аутофагия отвечает за удаление белков, поврежденных окислением, например, из мозга для восстановления его надлежащей функции [112, 113].
Во время старения митохондрии — основной источник АФК — часто подвергаются окислительному повреждению на уровне, превосходящем защитную способность антиоксидантного ответа.В таких случаях удаление поврежденных митохондрий посредством митофагии имеет решающее значение для смягчения пагубного воздействия на организм [114]. Более того, у C. elegans тесная связь между митофагией и митохондриальным биогенезом важна для обеспечения долголетия в условиях стресса [115]. Также было показано, что у мух сверхэкспрессия митофагического белка PARKIN увеличивает продолжительность жизни за счет увеличения митохондриального обмена [116]. Таким образом, митофагия выступает в качестве основного маркера повреждения, вызванного АФК, и играет важную роль в старении и различных возрастных расстройствах [117].
5. Связь ядра и митохондрий и важность митохондриального протеостаза
Повреждение ядерной ДНК индуцирует передачу сигналов от ядра к митохондриям (передача сигналов NM). Этот процесс играет жизненно важную роль в митохондриальном гомеостазе и старении. Ядерные белки (например, HIF-1 α , активатор пролифератора гамма-коактиватор-1 α (PGC-1 α ), белок O (FOXO) и семейство сиртуинов) вместе с повреждением ядерной ДНК Белки репарации могут влиять на целостность митохондрий и способствовать возрастным патологиям [118].Недавние исследования установили важную связь между никотинамидадениндинуклеотидом (NAD + ) и белками репарации ДНК в поддержании митохондриального метаболизма и увеличении продолжительности жизни [119].
Сиртуины, зависимые от NAD + деацетилазы, действуют как метаболические сенсоры, воспринимающие дисбаланс в соотношении NAD + / NADH. Ингибирование белков репарации ДНК, в частности белков поли (АДФ-рибозы) полимеразы (PARP-1 и PARP-2), потребляющих NAD + , увеличивает клеточные уровни NAD + [119].Высокие уровни NAD + впоследствии активируют сиртуины, которые, в свою очередь, способствуют более высокому содержанию митохондрий, увеличению расхода энергии и защите от метаболических заболеваний [119], в конечном итоге продлевая продолжительность жизни [120]. Кроме того, было показано, что активаторы сиртуина, такие как ресвератрол, способствуют долголетию [121, 122], вызывая эффекты ограничения калорийности (CR-) у C. elegans [123].
Однако и PARP, и сиртуины должны потреблять NAD + , чтобы работать.Большое количество PARP и сиртуинов может истощать клеточные уровни NAD + . Истощенные уровни NAD + приводят к инактивации сиртуинов и избыточной продукции ROS, что нарушает целостность митохондрий [124]. Более того, нарушения в активности сиртуинов дезактивируют несколько ферментов, включая PGC-1 α (гамма-коактиватор рецептора, активируемого пролифератором пероксисом 1 α ), факторы транскрипции вилочного блока O (FOXO), индуцируемый гипоксией фактор-1 α (HIF-1 α ) и AMP-активированная протеинкиназа (AMPK), которая модулирует выработку различных антиоксидантных ферментов, влияя на механизмы окислительной защиты [125].
Индуцированная повреждением ДНК передача сигналов NM через ось сиртуина PARP-NAD + может ускорить начало старения за счет нарушения целостности митохондрий. Таким образом, генетические или фармакологические вмешательства, направленные на белки или метаболиты, участвующие в передаче сигналов NM, могут потенциально способствовать долголетию. Например, у стареющих крыс лечение ингибитором PARP INO-1001 уменьшает сердечно-сосудистые нарушения [126], а лечение ингибитором PARP PJ34 улучшает сократительную функцию миокарда и восстанавливает функцию эндотелия [127].Более того, ингибирование PARP-1 может защищать от возрастной эндотелиальной дисфункции, потенциально за счет регулирования биодоступности NO через iNOS [128].
Однако положительная роль ингибирования PARP-1 в старении подвергается сомнению [129]. Например, PARP-1-нулевые мыши имеют сокращенную продолжительность жизни, более раннее начало старения и повышенную скорость спонтанного канцерогенеза по сравнению с мышами WT [130]. Одним из объяснений расхождений между исследованиями является двойная роль PARP: в то время как PARP способствует поддержанию геномной стабильности и увеличению продолжительности жизни, чрезмерная активность PARP истощает клеточный NAD + и запускает ядерный фактор- κ B- (NF- κ B). -) индуцированное воспаление, приводящее к быстрому старению и возрастным нарушениям [131].
Старение сопровождается снижением синтеза NAD + и увеличением потребления NAD + , что приводит к чистому уменьшению пула доступных NAD + (Рисунок 3). Снижение уровней NAD + приводит к возрастному снижению активности сиртуина 1 (SIRT1). Снижение активности SIRT1 влияет на функцию митохондрий, по крайней мере, с помощью двух механизмов: (1) снижение биогенеза, вторичное по отношению к снижению активности PGC1- α , и (2) нарушение функции митохондрий из-за снижения репликации и транскрипции митохондриальной ДНК [132, 133] .Таким образом, предполагается, что добавление NAD + или его предшественников способствует здоровому старению и долголетию [134–136].
Экспериментальные модели показали, что добавка NAD + полезна для поддержания углеводного обмена, сердечно-сосудистой функции, функции стволовых клеток и долголетия [137]. Более того, никотинамид предотвращает клеточное старение за счет снижения избыточной продукции АФК [138, 139]. Несколько клинических исследований на людях, проверяющих эффективность этого соединения, продолжаются [140].
Было показано, что улучшение продолжительности жизни C. elegans , опосредованное NAD + , включает ряд взаимосвязанных механизмов, которые включают (1) активацию гомолога сиртуина червя Sir-2.1, (2) ядерную транслокацию и активацию Фактор транскрипции FOXO daf-16 и (3) повышенная экспрессия антиоксидантных ферментов [141].
В модели на мышах обработка предшественником никотинамид рибозида (NR) NAD + задерживала старение мышечных и нервных стволовых клеток и увеличивала продолжительность жизни.Этот эффект, по-видимому, опосредован индукцией ответа митохондриального развернутого белка (UPRmt) [142]. Участие UPRmt в увеличении продолжительности жизни NAD + также было предложено в C. elegans [143].
UPRmt представляет собой форму ретроградной передачи сигналов, которая вносит вклад в обеспечение поддержания и функциональной целостности митохондриального протеома [144]. Накопление неправильно свернутых белков или несобранных комплексов в митохондриях сверх определенного порога приводит к измененному протеостазу, который может приводить к дисфункции органелл / клеток [145].Митохондрии передают это сообщение о бедствии в цитозоль и ядро через различные типы сигналов, и в ответ клетка вызывает набор ответов, включая выработку локализованных в митохондриях молекулярных шаперонов и протеаз, способствующих восстановлению гомеостаза органеллярных белков [91, 92 , 146, 147].
Было показано, что адаптивный путь, запускаемый сиртуин-зависимым UPRmt, который приводит к увеличению содержания и активности митохондриальных комплексов [143, 148], приводит к увеличению продолжительности жизни, по крайней мере, у мышей и мух [143, 142, 146] .Митохондриальная ретроградная передача сигналов в ядро через путь mTOR также увеличивает продолжительность жизни нормальных фибробластов человека, увеличивает потенциал митохондриальной мембраны, снижает уровень ROS и усиливает аутофагический поток [149]. АФК могут оказывать дополнительную нагрузку на систему контроля качества белков, поскольку сами белковые шапероны подвержены окислительному повреждению, что приводит к дальнейшему накоплению повреждений и ускоренному старению [4, 65].
В совокупности эти исследования устанавливают опосредованную ROS связь между митохондриями, ядром и протеостазом.
6. Роль АФК в нефармакологических стратегиях увеличения продолжительности жизни
6.1. Ограничение калорий (CR)
Термин «ограничение калорийности» обозначает снижение потребления энергии без недоедания и представляет собой наиболее эффективное и воспроизводимое диетическое вмешательство, которое, как известно, способствует здоровому старению и замедляет проявление возрастных расстройств у различных модельных организмов, включая дрожжи [150–153], нематоды [154, 155], плодовые мухи [156], мыши [157–159] и приматы [160].CR регулирует множество физиологических процессов, связанных со старением, включая метаболизм [161–165], окислительный стресс [166, 167], стабильность генома [168] и сигналы роста [169–171].
Для объяснения положительного эффекта ограничения калорийности были предложены четыре основные теории. Согласно гипотезе «ослабления окислительного повреждения», окислительное повреждение уменьшается во время ограничения калорий (CR) за счет снижения продукции активных форм кислорода и усиления защитных ферментов, что приводит к уменьшению повреждения ДНК и увеличению стабильности генома [168 , 172, 173].Гипотеза «глюкоза-инсулин» предполагает, что пониженные уровни циркулирующего инсулина и глюкозы, которые сопровождают CR, приводят к снижению роста и деления клеток, смещая ресурсы клетки в сторону поддержания и восстановления [172, 173]. Родственная гипотеза «инсулиноподобного фактора роста (IGF) 1» предполагает, что снижение уровней гормона роста и IGF-1 в ответ на CR способствует поддержанию и восстановлению [172, 173]. Наконец, гипотеза «стресс-адаптация» (или гормезис) предполагает, что CR способствует низкому уровню стресса, который вызывает перекрестную адаптацию к другим стрессовым факторам за счет увеличения уровней антиоксидантов и белков репарации ДНК [174].
Было предложено несколько молекулярных объяснений эффектов увеличения продолжительности жизни CR. Однако многое еще неизвестно о точном вкладе каждого пути в эффект продления жизни CR. Это понимание дополнительно осложняется обширным перекрестным взаимодействием между различными путями и тем фактом, что некоторые пути присутствуют в одних модельных организмах, но не присутствуют в других. Сложная сеть путей, которые участвуют в увеличении продолжительности жизни при ограничении калорийности, изображена на рисунке 4.
Два из наиболее изученных путей, предположительно участвующих в опосредованном CR продлении жизни, опосредованы ингибированием передачи сигналов инсулина / IGF-1 и инактивацией mTOR (механистическая мишень рапамицина). Оба считаются путями определения питательных веществ (инсулин для глюкозы и mTOR для аминокислот). Снижение циркулирующих уровней питательных веществ (аминокислот, глюкозы и даже холестерина), все из которых также воспринимаются mTOR, способствует снижению активности mTOR во время CR [175].Ингибирование mTOR приводит к SKN-1- / Nrf- и daf-16 — / FOXO-опосредованной активации защитных генов, что приводит к увеличению стрессоустойчивости и продолжительности жизни [176]. Кроме того, известно, что ингибирование mTOR вызывает аутофагию, которая играет важную роль в протеостазе во время старения [177, 178]. Эффект увеличения продолжительности жизни от ингибирования mTOR, генетически или химически, по-видимому, очень консервативен для разных модельных организмов [159, 179–181]. Инсулиновый путь опосредуется несколькими дополнительными ферментами, включая PI3K / Akt / Ras и транскрипционный фактор forkhead O (FOXO) [182–184].
Путь, опосредованный аденозинмонофосфат-активируемой протеинкиназой (AMPK), является третьим возможным CR-релевантным путем, который у некоторых организмов может пересекаться с путем mTOR. AMPK — это высококонсервативный сенсор повышенных уровней AMP и ADP, происходящих из-за истощения АТФ [185–187]. В общем, активация AMPK действует для поддержания запасов клеточной энергии, включая катаболические пути, которые производят АТФ, в основном за счет усиления окислительного метаболизма и митохондриального биогенеза, при этом отключая анаболические пути, которые потребляют АТФ.Важность AMPK в определении продолжительности жизни демонстрируется тем фактом, что лечение метформином, активатором AMPK, увеличивает продолжительность жизни C. elegans и короткоживущих линий мышей, склонных к раку [188–190].
Еще один важный путь — это путь, управляемый сиртуинами, активность которых увеличивается с CR. Ассоциация сиртуинов со сниженными уровнями окислительного стресса и повышенной антиоксидантной защитой была предложена для нескольких модельных организмов [191, 192], а также для людей, но точные молекулярные механизмы, лежащие в основе этой ассоциации, остаются неясными.Предполагается, что SIRT3 играет важную роль в усилении системы антиоксидантной защиты митохондрий глутатионом во время ограничения калорийности [193]. SIRT3-зависимая митохондриальная адаптация может также вносить вклад в замедление старения у млекопитающих [193].
Их роль в качестве посредников в благотворных эффектах, оказываемых ограничением калорийности, сделала сиртуины многообещающими фармакологическими мишенями для замедления старения и возрастных заболеваний [194]. Ресвератрол — это полифенольный антиоксидант, который содержится в красном вине и, как было показано, активирует сиртуины в нескольких организмах, включая человека [195, 196].Ресвератрол также является активатором AMPK, и эта активность также может способствовать положительному воздействию этого полифенола [197]. Предположительно, ресвератрол активирует механизмы антиоксидантной защиты и ослабляет выработку митохондриальных АФК за счет активации сиртуина. После приема ресвератрола наблюдались значительное снижение клеточной перекиси водорода [198–200], повышенная экспрессия MnSOD [195, 196] и повышенное содержание клеточного глутатиона [201]. Терапевтический потенциал ресвератрола был предметом интенсивных исследований в течение последнего десятилетия (например,г., [195–198]).
CR также снижает возрастное накопление окислительного повреждения за счет снижения митохондриального дыхания, мембранного потенциала и скорости продукции ROS [166, 167], хотя CR, по-видимому, оказывает лишь незначительное влияние на возрастные изменения. в митохондриальном протеоме [202]. CR также увеличивает митохондриальный биогенез через сигнальный путь PGC-1 α [203]. Более того, другие исследования также показали, что CR защищает от возрастных сосудистых нарушений, увеличивая биодоступность оксида азота (NO), снижая продукцию ROS, вызывая противовоспалительные реакции и предотвращая окислительное повреждение за счет активации элемента NRF-антиоксидантного ответа (ARE). сигнальный путь [204, 205].
Ограничение калорийности обычно подразумевает сокращение потребления пищи на 20-40% по сравнению с нормальным потреблением. Это довольно серьезное вмешательство, которое может иметь пагубные последствия [191]. Прерывистые или периодические диетические ограничения без хронического ограничения калорийности могут обеспечить значительное увеличение продолжительности жизни при минимизации побочных эффектов. Фактически, исследования на грызунах показали, что даже снижение потребления пищи на 10% может существенно повлиять на продолжительность жизни [206]. Sod — / — мыши демонстрируют повышенный уровень окислительного стресса, который, в свою очередь, приводит к сокращению продолжительности жизни. Было показано, что ограничение в питании (60% диеты ad libitum) увеличивает продолжительность жизни мышей Sod — / — на 30%, что делает ее похожей на продолжительность жизни контрольных мышей дикого типа, получавших ad libitum [207], за счет снижения перекисного окисления липидов в печени и головном мозге. Было обнаружено, что такое же диетическое вмешательство ослабляет возрастную атрофию мышц за счет снижения окислительного стресса у мышей даже при полном отсутствии ключевого антиоксидантного фермента CuZnSOD [208].
6.2. Exercise
Упражнения — еще одно эффективное нефармакологическое средство замедления негативных последствий старения. В нескольких исследованиях сообщалось о повышенной нагрузке O 2 в волокнах скелетных мышц [209, 210] и повышенных уровнях ROS [209, 211] во время упражнений в результате усиленного митохондриального дыхания, необходимого для выработки АТФ для мышечных сокращений. В то время как митохондриальное окислительное фосфорилирование является основным источником АФК, индуцированных упражнениями, ксантиноксидаза и эндотелиальная синтаза оксида азота (eNOS) также вносят вклад в образование АФК во время тренировок на выносливость [212] и упражнений на растяжку [40, 41].Регулярные физические упражнения связаны со снижением смертности и заболеваемости возрастными заболеваниями [213–215]. Следовательно, физические упражнения потенциально могут принести пользу пожилым людям за счет модуляции воспалительного и окислительно-восстановительного статуса, что может влиять на протеостаз, чувствительность к инсулину, состав тела (например, жировую ткань) и уровни гормонов [216].
Сообщалось о связанном со старением увеличении продукции АФК в клетках скелетных и сердечных мышц во время покоя и после тренировки [211, 217].На мышечном уровне возрастное повышение уровней АФК было связано с различными механизмами, такими как нарушение регуляции ETC из-за снижения активности цитохром-с-оксидазы и других ферментов [218] и нарушение митохондриальной мембраны из-за перекисного окисления липидов и ненасыщенности [44, 45].
Однако сообщалось и о противоречивых результатах. Исследование тканей скелетных и сердечных мышц старых крыс показало значительное увеличение антиоксидантных ферментов, таких как SOD, каталаза, GPX [47–50] и глутатион (GSH) [51, 132].Кроме того, мышцы, которые подвергаются хроническим упражнениям, демонстрируют более низкий окислительный стресс с точки зрения повреждения липидов, белков и ДНК как у людей, так и у модельных организмов [38, 49, 133]. Соответственно митохондрии, выделенные из тренированных мышечных клеток, показали более высокую устойчивость к окислению in vitro [55, 219]. Исследования также показывают повышение уровней SOD, GPX и GSH после тренировок на выносливость как у молодых, так и у пожилых людей [56, 57]. Эти результаты предполагают, что регулярные физические упражнения сопровождаются адаптацией клеток к окислительному стрессу, что, в свою очередь, оказывает положительное влияние, например, на иммунную систему [220].Эта идея резюмируется концепцией гормезиса. Гормезис можно определить как адаптивную реакцию, наблюдаемую у организмов, постоянно подвергающихся воздействию низкого или среднего уровня стресса. В этих условиях клетки развивают адаптивный ответ, включая повышенную экспрессию антиоксидантных генов, что, в свою очередь, делает их устойчивыми к множественным стрессорам [221, 222].
Индукция гормезиса контролируется окислительно-восстановительными сенсорными путями, которые после активации окислителями активируют антиоксидантную ферментативную систему [223].Например, интенсивные физические упражнения активируют митоген-активируемую протеинкиназу (MAPK) и редокс-сигнальные пути NF- κ B как у людей, так и у грызунов [60, 61]. Основными мишенями этих путей являются антиоксидантные ферменты, в том числе SOD, GPX и GSH, которые содержат сайты связывания NF- κ B и активаторного белка-1 (AP-1) в своих промоторах [62–65], а также реагирующие элементы. на различные стимулы, такие как провоспалительные цитокины, напряжение кислорода и ROS [66–68]. В скелетных мышцах другой важной горметической адаптацией к окислительному стрессу является увеличение митохондриальной массы и содержания белка [69, 70], особенно уровня цитохром с оксидазы.Цитохром с оксидаза контролирует поток электронов и образование супероксида в ЭТЦ [224]. Эти изменения повышают экспрессию PGC-1, который управляет биогенезом митохондрий в скелетных мышцах во время упражнений [225]. PGC-1 также связан со снижением окислительного стресса [226].
Было высказано предположение, что этот горметический ответ на окислительный стресс нарушается с возрастом скелетных мышц [218, 227]. Эта гипотеза подтверждается несколькими исследованиями, в которых сообщается о значительно более низкой экспрессии и активности NF- κ B в старых мышцах [217, 228, 229].Напротив, в других исследованиях сообщалось об неизменных [230] или даже более высоких [231] уровнях NF- κ B в состоянии покоя и о снижении активации пути MAPK после тренировки в старых мышцах грызунов и людей.
Хотя физические упражнения были предложены в качестве эффективных нефармакологических средств отсрочки негативного воздействия старения на функциональные и метаболические параметры [216], также хорошо известно, что регулярные энергичные упражнения могут иметь пагубные последствия, о чем свидетельствует повышенная восприимчивость элитные спортсмены к инфекциям [232].Этот эффект, по-видимому, по крайней мере частично, связан с пагубными последствиями длительного воздействия повышенной выработки АФК, связанного с интенсивными тренировками. Например, хроническое мышечное повреждение, распространенное не только у спортсменов, но и у пожилых людей, увеличивает выработку провоспалительных цитокинов, таких как фактор некроза опухоли- α (TNF- α ) и интерлейкин-6 (IL-6). ), которые дополнительно способствуют окислительному стрессу, который, в свою очередь, обостряет воспаление мышц, создавая порочный круг воспаления и окислительного повреждения [233].
Необходимы дополнительные исследования, чтобы разрешить противоречивые результаты относительно эффектов физических упражнений и гормезиса, вызванного физическими упражнениями, на статус окислительного стресса скелетных мышц и его прогрессирование на протяжении всей жизни.
7. АФК против старения: пусть бактерии возьмут верх
До недавнего времени считалось, что бактерии не подвергаются никаким событиям, эквивалентным процессу старения [234]. Однако за последнее десятилетие эта точка зрения изменилась. Впервые бактериальное старение было обнаружено у асимметрично делящегося Caulobacter crescentus [235, 236].У этих α -Протеобактерий деление клеток морфологически и функционально асимметрично. Эта асимметрия дает четкое различие между материнскими и дочерними клетками. Ackermann et al. [162] сообщили, что при множественных делениях время, необходимое материнской клетке для образования новой дочерней клетки, удваивается с 2,6 до 5 часов на цикл деления, процесс подобен репликативному старению у эукариот [235, 236]. Позже Стюарт и др. [237] продемонстрировали, что Escherichia coli также проявляют черты репликативного старения, несмотря на симметричное бинарное деление.Используя автоматическую покадровую микроскопию для изображения 8-10 циклов воспроизводства отдельных клеток, авторы заметили, что клетки со старым полюсом (материнские) показали пониженную приспособленность к росту (например, скорость роста) в течение последующих поколений по сравнению с их новым полюсом. (сестринские) клетки. Клетки со старым полюсом снизили образование потомства и увеличили частоту гибели клеток. Примерно через 100 делений клетки со старым полюсом перестали расти [237].
Последующие исследования показали, что аналогичные процессы происходят у Bacillus subtilis [238] и Mycobacterium spp.[239] (Рисунок 5). Эти наблюдения подтверждают, что старение у бактерий является более общим явлением, чем считалось ранее, которое влияет не только на микробы с разной морфологией внутри линии от матери к дочери, но и на тех, у которых асимметрия роста наблюдается в потомстве на функциональном / молекулярном уровне. .
7.1. Старение и условное старение
Во время стационарной фазы в результате ограничения питательных веществ клетки E. coli входят в уникальное состояние, известное как условное старение [240].После того, как бактерии стареют, они постоянно теряют способность к культивированию и не могут возобновить рост, даже когда питательные вещества снова становятся доступными. Эта особенность делает условное старение очень похожим на старение соматических клеток человека [100–103] и репликативную продолжительность жизни дрожжей ( S. cerevisiae ), которая обычно используется для моделирования процесса старения митотических тканей у высших организмов [241] .
Наблюдаемая функциональная асимметрия в бактериальном делении, о которой первоначально сообщили Stewart et al.[237], был связан с асимметричной сегрегацией поврежденных клеточных компонентов (например, белковых агрегатов) [242, 243], процесс также присутствует у эукариот [244–246]. Асимметричная агрегация повреждений белков, по-видимому, является активным процессом у дрожжей [87]. У бактерий этот процесс, по-видимому, в основном пассивен и управляется молекулярным скоплением [247].
Пакетные культуры E. coli , подвергнутые остановке роста, вызванной голоданием, демонстрируют заметно более высокие нагрузки поврежденных (карбонилированных) белков [248], что также присутствует у стареющих эукариот [249, 250].Однако эта нагрузка не распределяется среди населения равномерно. Интересно, что клетки с низкой карбонильной нагрузкой оставались репродуктивно компетентными, тогда как клетки с высокой карбонильной нагрузкой были генетически мертвы (то есть не могли культивироваться). Запрограммирована ли эта вызванная голоданием гетерогенность карбонилирования и приспособленности и является ли она результатом сегрегации повреждений во время цитокинеза, не выяснено. Старение бактериальных клеток, вызванное другими внешними стимулами, включая УФА-излучение, также связано с накоплением белковых карбонилов в результате окислительного повреждения [251–253] (рис. 6).
Зависимое от времени накопление белковых карбонилов наблюдалось во время стационарной фазы у E. coli [254]. Активности, которые способствуют окислению белка во время стационарной фазы, показаны на рисунке 7. Учитывая, что одним из критериев старения является увеличение смертности с течением времени [255], это зависящее от времени накопление карбонилов белка является убедительным аргументом в пользу того, что прокариоты, такие как E. coli , возраст. Некоторые белки, такие как ферменты цикла трикарбоновых кислот (ТСА), по-видимому, особенно чувствительны к карбонилированию [256].Интересно, что клетки, лишенные активности SOD-1, обнаруживают большее количество карбонилирования белка и быстрее теряют жизнеспособность в стационарной фазе [248]. Более того, популяции в стационарной фазе, инкубированные в отсутствие кислорода, имеют значительно увеличенную продолжительность жизни по сравнению с аналогами, выращенными в присутствии кислорода [254]. Эти наблюдения подчеркивают участие АФК и окислительного стресса в старении, связанном со стационарной фазой.
В стационарной фазе и в стрессовых условиях окисление специфических белков в E.coli . Эти белки включают DnaK (шаперон Hsp70), ДНК-связывающий белок H-NS, универсальный стрессовый белок A (UspA), факторы элонгации EF-Tu и EF-G, глутамин синтетазу, глутаматсинтазу и аконитазу [254, 256, с. 257]. Интересно, что некоторые из этих белков также карбонилируются в дрожжевых клетках при окислительном стрессе [258], у стареющих мух [259, 260] и в человеческом мозге людей с болезнью Альцгеймера [261]. Эти наблюдения предполагают, что неконтролируемое окислительное повреждение в форме карбонилирования белка могло быть ближайшей причиной старения среди E.coli [248]. Однако прямых доказательств этой гипотезы нет.
При остановке роста, стационарная фаза E. coli развивает устойчивость к тепловому и окислительному стрессу, явление, известное как индуцированная стазисом перекрестная защита [262]. Клетки, лишенные углерода или азота, заметно более устойчивы к тепловому шоку и окислительному стрессу, чем пролиферирующие клетки [262, 263]. Связь между стрессоустойчивостью и продолжительностью жизни была описана на нескольких модельных эукариотических организмах [264–267].Эти наблюдения указывают на то, что может существовать эволюционно сохраняющийся механизм, направляющий ресурсы от воспроизводства к поддерживающим и защитным функциям [268]. Подобно эукариотам, способность клеток подавлять АФК может играть роль в определении продолжительности жизни бактерий [248].
Однако, как и в случае с эукариотами, существуют противоречивые результаты относительно вклада АФК в бактериальное старение. Например, репродуктивно арестованные популяции E. coli имеют повышенные уровни белков окислительной защиты и повышенную устойчивость популяции к внешним окислительным стрессам [138, 139].Однако в этих популяциях также обнаруживаются более высокие уровни поврежденных белков [254, 256]. Кроме того, не наблюдается строгой корреляции между дыхательной активностью, окислением белков и продолжительностью жизни бактерий E. coli с задержкой роста [269]. Подобные результаты наблюдались в дрожжевых клетках с задержкой роста G0 [270].
Первые гены, индуцированные после остановки роста у бактерий, играют роль в противодействии старению и гибели, вызванным стазисом [262, 271]. Многие из этих генов кодируют белки, которые защищают клетку от внешних стрессов, таких как тепло, оксиданты и осмотическая нагрузка, что может объяснить индуцированную стазисом перекрестную защиту [262].Перекрестная защита основана на сигма-факторе Sigma-S [272]. Не только при голодании, но и в условиях общего стресса, фактор транскрипции Sigma-S накапливает, связывает и направляет РНК-полимеразы к более чем 50 специфическим генам [272]. E. coli мутанты, лишенные Sigma-S, имеют повышенные уровни белков с окислительным повреждением [254, 256] и ускоренное старение во время остановки роста [272]. У Salmonella sp. Как Sigma-S, так и Sigma-E необходимы для защиты от окислительного повреждения в стационарной фазе, а мутанты, лишенные Sigma-E, имеют пониженную выживаемость и повышенную восприимчивость к окислительному стрессу [273].Однако в условиях анаэробной стационарной фазы выживаемость полностью сохраняется [273], что указывает на то, что окислительное повреждение является основным механизмом, с помощью которого снижается жизнеспособность микробов во время недостатка питательных веществ. Интересно, что члены регулона Sigma-S включают разнообразный набор белков, функции которых перекрывают функции регулируемых FOXO- / daf-16 генов долголетия у C. elegans [142, 147, 148]. Таким образом, функционально сходные сигнальные пути, по-видимому, регулируют устойчивость к стрессу, защиту от повреждения белков и продолжительность жизни у эукариот и прокариот.Эти пути имеют решающее значение для выживания в периоды голодания. Возможно, они эволюционно сохранились на разных ветвях древа жизни, потому что они увеличивали способность клетки к поддержанию. Со временем они также могут стать решающими в замедлении старения многоклеточных организмов [143, 149].
7.2. Генетические детерминанты старения и старения бактерий
Литература, посвященная изучению генов, увеличивающих выживаемость в стационарной фазе у бактерий, недостаточна.Однако сообщалось о нескольких мутантных штаммах, которые выживают дольше, чем WT. Было обнаружено, что RssB, который регулирует стабильность сигма-фактора Sigma-S, играет ключевую роль в выживании E. coli , потенциально за счет повышения устойчивости клетки к спонтанным эндогенным стрессам [274].
Совсем недавно полногеномный скрининг мутантов E. coli с фенотипом продолжительной стационарной фазы выживания выявил три штамма, которые жили дольше, чем WT [275].Один из штаммов, Δ sdhA (субъединица А сукцинатдегидрогеназы), проявил повышенную стрессоустойчивость и увеличил продолжительность жизни. Сукцинатдегидрогеназа представляет собой тетрамерный белковый комплекс, который катализирует превращение сукцината в фумарат в цикле TCA [276]. Субъединица A, ферментативно активная часть комплекса, является хорошо известным источником супероксида в ETC E. coli [277]. Предположительно, когда этот фермент отсутствует, скорость производства супероксида снижается, что увеличивает выживаемость в стационарной фазе [275].
Двумя другими мутантами, демонстрирующими длительную выживаемость в стационарной фазе, были Δ lipA (липоилсинтаза) и Δ lpdA (дигидролипоилдегидрогеназа) [275]. Авторы объясняют увеличенную продолжительность жизни этих двух мутантов сниженным потреблением кислорода по сравнению с WT, что, в свою очередь, увеличивает экспрессию фактора транскрипции гипоксии ArcA [278]. ArcA подавляет экспрессию генов цикла TCA, таких как цитрат-синтаза ( gltA ), и активирует экспрессию генов, необходимых для выработки энергии в условиях ограниченного кислорода, увеличивая выживаемость в стационарной фазе [279].Эти наблюдения предполагают, что увеличенная продолжительность жизни, наблюдаемая у этих мутантов, связана с индукцией физиологического состояния, обычно связанного с гипоксическими состояниями. Эти результаты согласуются с ролью HIF-1 α в модуляции продолжительности жизни у высших организмов [37, 162, 163]. Фактически, ArcA можно рассматривать как функциональный гомолог HIF-1 α , хотя эти два белка не обладают значительным сходством последовательностей. Это функциональное сходство указывает на адаптивный ответ к условиям с ограничением кислорода как на эволюционно законсервированный механизм, который может увеличивать продолжительность жизни.
Учитывая сохранение ключевых фенотипов, связанных с возрастными макромолекулярными повреждениями, и роль генов, контролирующих гипоксический ответ как у бактерий, так и у высших эукариот, в увеличении продолжительности жизни, разумно предположить, что наиболее фундаментальные механизмы старения могут сохраняться. на всех уровнях жизни. Будущие исследования помогут выяснить, какие молекулярные процессы, лежащие в основе старения, похожи у бактерий и эукариот. Результаты этих исследований могут открыть возможность использования E.coli как модельный организм старения, на котором можно легко проверить конкретные молекулярные механизмы и эволюционные теории.
8. Выводы и перспективы на будущее
Прогрессирующая потеря митохондриальной функции является постоянным и сохраняющимся признаком старения, который влияет как на клеточный гомеостаз, так и на здоровье организма [134, 135]. Хотя АФК способствуют старению, они также играют решающую роль в передаче сигналов и развитии клеток, тем самым играя полезную роль. Митохондриальная теория старения предлагает примирительную перспективу двойной роли ROS в процессе старения, включая два важных адаптивных ответа: (1) UPRmt-опосредованную ретроградную передачу сигналов от митохондрий к ядру для регулирования старения и (2) ROS-опосредованную передачу сигналов. адаптивный ответ для активации системы антиоксидантной защиты клетки.Вмешательства, направленные на любой из этих двух адаптивных путей, можно рассматривать как потенциальные цели для терапии против старения и увеличения продолжительности жизни.
Поскольку накопление окислительных повреждений на протяжении всей жизни является основной причиной старения, генетические или фармакологические вмешательства, направленные на восстановление или устранение окислительных повреждений, сами по себе также имеют значительный терапевтический потенциал. Однако необходимы дальнейшие исследования на людях и нечеловеческих приматах, чтобы понять клиническую значимость потенциальных генетических, фармакологических и нефармакологических вмешательств.
Наблюдение за тем, что некоторые из процессов, характеризующих старение эукариот, также можно наблюдать у бактерий, подчеркивает потенциал бактерий в качестве простого модельного организма для изучения старения и связанных с возрастом механизмов. Эти поддающиеся обработке модели могут оказать решающую помощь в поисках генетических, молекулярных и биохимических процессов, лежащих в основе старения и возрастных заболеваний.
Раскрытие информации
Спонсоры не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.Текущий адрес Санчари Синха — независимый научный писатель, Калькутта, Индия.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов в отношении содержания этой статьи.
Благодарности
Благодарности анонимному рецензенту за подробный анализ и полезную критику предыдущей версии этой рукописи. Авторы также хотели бы поблагодарить доктора Натана Сусника (Ганноверская медицинская школа, Ганновер, Германия) за его проницательные комментарии и вдумчивые советы.Авторы выражают благодарность Фонду исследований AXA (http://www.axa-research.org/) за поддержку.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie.Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
генов | Бесплатный полнотекстовый | Роль элонгатор-зависимых путей модификации тРНК в нейродегенерации и раке
1. Регуляция эпитранскриптомной трансферной РНК
Помимо четырех стандартных нуклеозидов, клеточная РНК содержит широкий спектр посттранскрипционных модификаций, которые могут оказывать значительное влияние на ее функцию [1]. Полный набор модификаций РНК клетки называется эпитранскриптомом, и его динамические изменения, как полагают, представляют стратегию модуляции функции РНК с участием «писателей» (модификаторов), «стирателей» (демодификаторов) и «читателей», белков, специфически распознающих РНК. модификации [2,3].Среди различных типов РНК, подверженных эпитранскриптомным изменениям, транспортная РНК (тРНК) несет в себе самые многочисленные и химически разнообразные модификации (рис. 1). Хотя изначально считалось, что они являются конститутивно модифицированными, к настоящему времени известны многочисленные примеры, демонстрирующие динамические изменения в паттернах модификации тРНК в ответ на изменение окружающей среды или условий [4,5,6,7,8,9,10,11,12,13]. Один из них включает пишущую метилазу и стирающую деметилазу, как это наблюдается в эпитранскриптомной регуляции трансляции матричной РНК (мРНК) [10].В этом случае ALKBh2 млекопитающих была идентифицирована как деметилаза, удаляющая метильную группу в 1-метиладенозине (m 1 A), которая обнаруживается в инициаторных и элонгирующих тРНК и вводится TRMT6 (субъединица, связывающая субстрат) и TRMT61 (каталитическая субъединица). ) [10]. Было продемонстрировано, что эпитранскриптомная модуляция m 1 Наличие А в обоих типах тРНК (инициатор и элонгатор) регулирует функцию тРНК в инициации и удлинении трансляции динамически в ответ на доступность глюкозы [10].Важно отметить, что отсутствие белка ластика вызывает эмбриональную летальность или нейронные дефекты в модельной системе мышей, что указывает на прямую или косвенную роль эпитранскриптомных изменений в тРНК млекопитающих в предотвращении заболевания [10]. Несмотря на тот факт, что m 1 A представляет собой Это единственная известная до сих пор модификация тРНК, которая активно удаляется белком-стирателем, множество дополнительных модификаций изменяется в их относительном количестве в ответ на изменение окружающей среды или физиологических условий.Эти триггеры включают повышенную температуру, окислительный стресс или доступность питательных веществ, необходимых для синтеза модификаций, например, серы, бикарбоната, квиена или таурина [4,5,6,9,11,13]. В связи с растущим объемом доказательств динамической, а не статической природы модификаций тРНК, следует рассматривать дополнительные примеры эпитранскриптомной регуляции функции тРНК.2. Модификации РНК переноса, связанные с заболеванием
Для того, чтобы тРНК функционировала в качестве адапторной молекулы при трансляции, необходимо ее складывание в l-образную структуру [1].Структура характеризуется третичным спариванием оснований и наличием оснований, образованных внутримолекулярным спариванием оснований в дополнение к одноцепочечным петлям. Ряд посттранскрипционных модификаций в неспаренных участках петли тРНК вносят вклад в структуру тРНК, предотвращая каноническое спаривание оснований [15]. Не только цитоплазматическая, но и митохондриальная тРНК подвержена динамической модификации ферментами, кодируемыми в ядре [9,11]. Различные заболевания человека связаны с мутациями в генах, необходимых для внесения модификаций [16,17,18,19,20] как в цитоплазматические, так и в митохондриальные тРНК (рисунок 1 и таблица 1).Заболевания, связанные с дефектами генов модификации тРНК человека, включают различные неврологические синдромы, а также метаболические и респираторные дисфункции (таблица 1). Кроме того, повышенная регуляция различных генов модификации тРНК наблюдается при различных типах рака (таблица 1), что указывает на повышенную потребность в активности модификаторов тРНК в различных типах опухолевых клеток. быть наиболее распространенным. К ним относятся умственная отсталость, семейная дизавтономия (ФД), эпилепсия и другие синдромы.Интересно, что различные мутации, которые, как известно, вызывают дефекты в различных модификациях тРНК, таких как 5-метилцитозин (m 5 C), 5-метоксикарбонилметлуридин (mcm 5 U), 5-метоксикарбонилметил-2-тиуридин (mcm 5 s 2 U), 5-карбамоилметилуридин (ncm 5 U), псевдоуридин (Ψ) в положениях 38/39 и 13/35, 1-метилгуанозин (m 1 G), инозин (I) и N2, N2- диметилгуанозин (m 2,2 G) связаны с умственной отсталостью у людей (таблица 1), что позволяет предположить, что нормальная функция и развитие мозга особенно зависят от наличия этих модификаций.Принимая во внимание положение критических модификаций внутри тРНК (рис. 1), примечательной особенностью является то, что многие из релевантных для заболевания модификаций тРНК естественным образом обнаруживаются внутри петли антикодона и, более конкретно, в положении колебания (34) антикодона. Дефекты модификации основания колебания в митохондриальных тРНК также связаны с различными миопатиями и могут также вызывать неврологические симптомы (Таблица 1) [19,20,37], подчеркивая центральную роль модификаций основания колебания для нормального функционирования тРНК в различных клеточных компартментах.Известно, что модификации в положении вобуляции позволяют расширить возможности спаривания оснований (инозин) [68] или улучшить точность трансляции и удлинение за счет оптимизации скорости трансляции кодонов (mcm 5 s 2 U) [69,70,71, 72,73,74]. Колебательные уридиновые модификации типа xm 5 U (mcm 5 U, mcm 5 s 2 U и ncm 5 U; Рисунок 1B) относятся к наиболее сложным модификациям тРНК с точки зрения вовлеченных генов ( Таблица 2) и путей, а также в отношении посттрансляционной модификации самих комплексов модификаторов тРНК [66].Множественные корреляции дефектов xm 5 U (Таблица 1) с заболеваниями человека указывают на то, что это семейство модификаций имеет выдающееся значение для здоровья и развития человека (Рисунок 2).3. Зависимые от элонгатора модификации РНК переноса
В антикодонах субнабора тРНК модификации уридинов вобблирования (U 34 ) требуют активности комплекса элонгатора (последние обзоры см. [18,75,76]). Целевые тРНК, получающие элонгатор-зависимые модификации, представляют собой тРНК Lys UUU , тРНК Gln UUG , тРНК Glu UUC , тРНК Arg UCU , тРНК3 GlyCC
3 GlyCC
3 UGA , тРНК Pro UGG , тРНК Thr UGU , тРНК Ala UGC и тРНК Leu UAA [77,78].Элонгатор — это мультисубъединичный (Elp1-Elp6) белковый комплекс, который был первоначально выделен из дрожжей в сочетании с удлиняющим голоферментом РНК-полимеразы II [79,80] и, следовательно, вовлечен в транскрипцию мРНК, а не трансляцию [78,81,82] Однако теперь известно, что Elongator собирается в активный голо-додекамерный комплекс [(Elp1-Elp6) 2 ], который связывает тРНК с разными субъединицами [83,84,85,86,87,88]. Соответственно, Elongator действует как модификатор тРНК в консервативном пути, который обнаруживается во всех трех сферах жизни и, как было показано, может функционально обмениваться между эукариотическими модельными организмами [86,89,90,91,92,93].В дрожжах комплекс присоединяется к U 34 из одиннадцати различных видов тРНК 5-карбоксиметил (см 5 ) групп [77,78], которые вместе с дополнительными ферментативными каскадами модификаторов U 34 могут быть дополнительно преобразованы до еще более сложных модификаций, включая (но не ограничиваясь ими) ncm 5 , mcm 5 или mcm 5 s 2 (см. рисунок 1B, [77,78]). Формирование боковой цепи cm 5 непосредственно катализируется Elp3, каталитической основной субъединицей Elongator, которая несет связывающие домены для тРНК и ацетил-CoA и радикальный мотив SAM (S-аденозил-метионин), который координирует металлоорганический [4Fe – 4S ] Кластер SAM , как было показано, имеет решающее значение для модификации U 34 in vivo [78,81,89,94].В поддержку этого предположения было показано, что архей Elp3 из Methanocaldococcus infernus модифицирует U 34 в петле антикодона синтетических субстратов тРНК Arg UCU in vitro в радикальной SAM-зависимой реакции, содержащей донор электронов (Na 2 S 2 O 4 ), SAM и ацетил-КоА [93]. Более того, сложное понимание основы и механизма способности модификации тРНК элонгатора было недавно получено благодаря элегантной структурно-функциональной работе над прокариотическим Elp3 из Dehalococcoides mccartyi и голо-элонгатором из дрожжей Saccharomyces cerevisiae [86,87,88,95,96, 97].Помимо биохимического выделения элонгатора (см. Выше), генетический скрининг, направленный на изоляцию дрожжевых мутантов, устойчивых к антикодонному расщеплению зимоцином, риботоксином тРНКазы, зависящим от модификации U 34 , также сыграл важную роль в идентификации субъединиц элонгатора и вспомогательных белков с ролями связанные с модификацией и регулированием U 34 [78,98,99,100,101,102,103,104]. Вместе с другими стратегиями, разработанными многими независимыми исследовательскими лабораториями, эти подходы в совокупности привели к идентификации комплекса метилазы колебания U 34 (Trm9 • Trm112) [101,105,106,107] и компонентов пути (Nfs1, Tum1, Uba4, Urm1, Ncs2, Ncs6) для связанного с элонгатором тиолирования U 34 [99,101,108,109,110,111,112,113,114,115,116].Метилтрансферазная (МТазная) активность Trm9 • Trm112, по-видимому, зависит от активности элонгатора для согласованного образования mcm 5 и mcm 5 s 2 в дрожжах и связанных модификаций гомологами метилазы колебания (ALKBH8 • TRMT112) в дрожжах. высшие эукариотические системы [117,118,119,120]. Интересно, что Trm112 не только способствует активности МТазы каталитической субъединицы Trm9, но также действует как активирующий узел или платформа для еще трех SAM-зависимых МТаз, модифицирующих рибосомную РНК (Bud23), тРНК (Trm11) или фактор терминации трансляции eRF1 (Mtq2) [ 121,122].В совокупности это иллюстрирует важность метилтрансфераз для трансляции мРНК и биосинтеза белка de novo. Что касается тиолирования U 34 и образования модификации mcm 5 s 2 , было показано, что включение S в U 34 субстратной тРНК тиолазы дрожжевого колебания (Ncs2 • Ncs6 aka CTU1 • CTU2, см. Рисунок 1) требует консервативной S-релейной системы (Nfs1, Tum1, Uba4) для S-активации, S-мобилизации и S-переноса на Urm1, a белок-модификатор, связанный с убиквитином [109,110,111,112,113,114].Urm1 уникален тем, что сочетает свойства, типичные для прокариотических S-носителей, с эукариотическими белками убиквитина [123,124]. В соответствии с этим представлением, Urm1 может действовать как неканонический, лизин-направленный модификатор белка в пути, известном как урмилирование белка, и как донор серы для тиолирования U 34 [112,113,114,125]. Обе роли строго зависят от серы и могут обмениваться среди эукариот [115,116,126] и основаны на Urm1-подобных белках из архей и бактерий [127,128]; они кажутся сохраненными и важными во всех сферах жизни.В соответствии с этим их инактивация вызывает вызванные стрессом дефекты роста у микробов, недоразвитие органов у растений и, что удивительно, летальность у мух [108,113,124,129,130,131]. В отличие от активации убиквитина и конъюгации с помощью обычных каскадов ферментов E1-E2-E3, активность E2 / E3 для Urm1 неизвестна, а активация Urm1 его E1-подобным ферментом Uba4 приводит к C-концевому тиокарбоксилированию (Urm1-COSH ), который имеет решающее значение как для урмилирования, так и для тиолирования тРНК [113,115,123,132,133].Образование Urm1-COSH очень похоже, если не идентично E1-подобной (MoeB или ThiF) активации бактериальных белков-носителей S (MoaD или ThiS), которые (а не участвуют в конъюгации белков) исключительно отдают серу для синтеза молибдоптерина. или кофакторы тиамина [134,135]. Таким образом, помимо его сходства с эукариотическими убиквитин-подобными белками, Urm1 действительно относится к прокариотическим белкам-носителям S, поэтому белок был придуман как молекулярное ископаемое на перекрестке модификаций белка и РНК [123].Для его двойной функциональности требуется десульфураза Nfs1, которая мобилизует серу из цистеина для прямого S-переноса на Uba4 [110,111] или косвенно через серотрансферазу Tum1 [109,112,113,114]. Uba4 снабжен MoeB-подобными (MoeBD, см. Выше) доменами типа роданезы (RHD), которые несут тиол-активные цистеины [113, 133]. S-перенос на один в RHD приводит к персульфиду, который после аденилирования Urm1 с помощью MoeBD, вероятно, образует ацилдисульфид с модификатором Urm1 [132,133,136]. После восстановительного расщепления этой связи Urm1-COSH высвобождается [137], чтобы быть способным действовать при урмилировании или отдавать активированные частицы серы для S-вставки в тРНК с помощью тиолазы Ncs2 • Ncs6 [115,133].Недавно в эукариотических Ncs6 и TtuA (родственная тиоуридинсинтетаза из термофильных бактерий и архей) были идентифицированы кластеры [3Fe-4S] и [4Fe-4S], соответственно, которые, по-видимому, участвуют (прямо или косвенно) в тио-модификации реакция, необходимая для образования s 2 U. Вместе с выводами о том, что тиолирование тРНК, по-видимому, не требуется для урмилирования белка, и наоборот, две ветви пути URM1 — хотя и механически связаны посредством активации серы — по-видимому, функционально отделены от них, а не зависят от них. друг друга [115].Таким образом, ранее предложенная концепция, согласно которой поток серы от Urm1-COSH к тиолированию тРНК может контролироваться с помощью урмилирования [138], кажется менее вероятным в настоящее время. Однако в этом контексте следует отметить, что человеческий URM1 (hURM1) и Urm1-подобные белки (SAMP, TtuB), как было показано, образуют урмилированные конъюгаты с человеческими и прокариотическими ортологами дрожжевой тиолазы (CTU1, CTU2, NcsA, TtuA) [ 125 127 128]. Означает ли это, что S-перенос (через Urm1-COSH) для тиолирования тРНК может включать прямое урмилирование субъединиц тиолазы, неизвестно, но привлекательно для поддержки варианта взаимозависимости между двумя ветвями пути URM1, урмилирования белка и тиолирования U 34 .Хотя S-донорная роль Urm1 для тиолирования тРНК была продемонстрирована in vitro [137], мы не знаем о переносе серы во время лизин-направленного урмилирования белковых мишеней (включая компоненты тиолазы, см. Выше) в дрожжах или других модельных организмах. Тем не менее на клеточных линиях человека было показано, что IKAP (гомолог дрожжевой элонгаторной субъединицы Elp1) урмилировался в условиях окислительного стресса [116]. Также были предложены дополнительные белки-мишени умилирования, включая субъединицы тиолазы человека CTU1 и CTU2 (см. Выше) [125].Поскольку актуальность урмилирования IKAP для функции модификации Elongator U 34 в человеческих клетках не изучена, и функциональная связь (если таковая имеется) между урмилированием IKAP / Elp1 и тиолазой (CTU1 • CTU2) в настоящее время неясна, эти интригующие явления еще предстоит разрешить в будущем.
5. Роль агрегации белков в нейродегенерации
Использование рибосомного профилирования, отсутствие mcm 5 s 2 U в дрожжевой тРНК Gln UUG и тРНК Lys UUU было показано, что приводило к замедление трансляции по родственным кодонам глутамина и лизина [70].Этот дефект идет рука об руку с накоплением клеточных белковых агрегатов [70]. В то время как прямой механизм образования агрегатов еще не выяснен, поразительным наблюдением было то, что существует большое перекрытие между агрегатами, вызванными гипомодификацией тРНК, и теми, которые образуются в отсутствие функционального шаперонного комплекса, ассоциированного с рибосомами (ssb1 / ssb2) [70,142]. Это может указывать на то, что замедление рибосом нарушает складывание возникающих полипептидов. Генетические подходы к почкующимся дрожжам дополнительно подтверждают, что образование белковых агрегатов в отсутствие различных важных модификаций тРНК представляет собой ключевой пусковой механизм плейотропных клеточных дефектов в клеточной полярности, морфогенезе и сегрегации ядер во время клеточного деления [143].Важно отметить, что агрегация белка в отсутствие модификации U 34 не ограничивается модельной системой дрожжей, но также наблюдалась у нематод, мышей и клеток человека [27,32,33,70,144,145]. Кроме того, отсутствие митохондриально-специфических таурин-производных модификаций колеблющегося уридина вызывает агрегацию неправильно нацеленных митохондриальных белков [146]. Интересно, что агрегация белков также является признаком различных нейродегенеративных заболеваний и, следовательно, может представлять функциональную связь между дефектами модификации тРНК (затрагивающими mcm 5 s 2 U и другие модификации) и нейродегенеративными заболеваниями или заболеваниями нервного развития.В подтверждение этого уже было продемонстрировано, что условный нокаут ELP3 у мышей индуцировал развернутый белковый ответ (UPR) и вызывал уменьшение количества нейронов кортикальных проекций, что приводило к дефектам развития нервной системы и микроцефалии [144]. В то время как индукция UPR у мутантов, модифицирующих тРНК, по-видимому, ограничивается системами высших эукариотических клеток, агрегация эндогенных цитоплазматических белков наблюдается как в клетках млекопитающих, так и в клетках дрожжей [27,32,33,70,144,145,147]. Поскольку мутации в ELP3 также связаны с фатальным исходом. Также анализировали дегенеративное заболевание двигательных нейронов БАС (см. выше и Таблицу 1), агрегацию белков в мышиной линии двигательных нейронов.После истощения ELP3 из этой клеточной линии наблюдалась агрегация белка, включая релевантную для ALS мутантную форму SOD1 [27]. Кроме того, несколько линий доказательств указывают на нейродегенеративные эффекты мутации ELP6, вызывающие атаксийные синдромы на модели мышей (см. Выше), которые сопровождаются агрегацией белков [141]. Кроме того, подавление либо ELP3, либо CTU2 (s 2 U) в клетках меланомы человека приводило к индукции эндогенных белковых агрегатов [32], предполагая, что влияние дефектов тиолирования элонгатора и тРНК на растворимость белка является общим и весьма вероятным заболеванием уместным.В последнем случае также было замечено, что ключевой фактор, перепрограммирующий экспрессию генов раковых клеток (см. Ниже), зависит от модификации уридина колебанием для эффективной трансляции и присутствует в эндогенных агрегатах [32]. Это наблюдение согласуется с предположением, что снижение скорости трансляции кодонов в отсутствие mcm 5 s 2 U вызывает дефекты сворачивания белка и агрегацию растущего полипептида. Обнаружение того, что агрегация белка может быть также вызвана у дрожжей дефектами в других модификациях, таких как Ψ 38/39 или t 6 A [143,148], и что те же самые дефекты в клетках человека вызывают связанные с ней нейродегенеративные синдромы / синдромы нервного развития [ 41,47,60] обеспечивают твердую поддержку предположению, что агрегация белков может представлять собой важный пусковой механизм нейродегенеративного заболевания, коррелированного с мутациями в модификаторах тРНК.6. U
34 Модификация при раке Помимо их роли в предотвращении нейродегенерации, ферменты, вводящие модификации тРНК U 34 , также были определены как ключевые факторы для поддержания метастазирования рака груди и мочевого пузыря и выживания клеток злокачественной меланомы [ 32,33,149]. В течение некоторого времени было известно, что гены в путях репарации повреждений ДНК обнаруживают определенное кодонное смещение [130, 150] и что потеря элонгатора или родственных путей (например, Uba4, Urm1, см. Выше) может вызывать повреждение ДНК [151].Эти знания недавно были расширены, показывая, что одни и те же пути являются решающими детерминантами выживания устойчивых к терапии клеток меланомы. В частности, мутация BRAF V600E является наиболее распространенной среди пациентов с меланомой человека и отвечает за устойчивость к таргетной терапии. ELP1, ELP3, CTU1 и CTU2 сильно активируются в клетках BRAF V600E , и инактивация ELP3 нарушает развитие меланомы BRAF V600E в модели рыбок данио [32].Более того, было показано, что комплекс Elongator способствует гликолизу в клетках меланомы посредством прямой кодон-зависимой регуляции трансляции мРНК фактора 1, индуцированного гипоксией (HIF1A), и поддержания высоких уровней белка HIF1α, обеспечивающего сильную устойчивость к антибиотикам. -BRAF терапия [32]. В предыдущем исследовании было показано, что миграция и онкогенность клеток, происходящих от меланомы, значительно снижается при истощении ELP1, ELP3, ELP5 или ELP6 в клетках меланомы [31]. В другом исследовании было показано, что ELP3 активируется в клетках гепатоцеллюлярной карциномы человека (HCC), что хорошо коррелирует с фосфорилированием протеинкиназы B (AKT) [152].Миграция и инвазия клеток HCC, опосредованная элонгатором, дополнительно стимулируется индуцированной экспрессией MMP-2 и MMP-9 через сигнальный путь PI3K (фосфоинозитид-3-киназа) / AKT. Было также показано, что ELP3 управляет Wnt-зависимой инициацией и регенерацией опухолей в кишечнике, поддерживая субпопуляцию клеток, экспрессирующих клетки LGR5 и SOX9 [153]. Кроме того, генетическое устранение ELP3 сильно нарушает инвазию и образование метастазов в модельной системе инвазивного рака груди. Более подробно, ELP3 и CTU1 • CTU2 активируются при инвазивном раке груди человека и поддерживают клеточную инвазию за счет трансляции онкопротеина DEK, который впоследствии способствует трансляции проинвазивного фактора транскрипции LEF1, зависящей от IRES (внутренний сайт входа в рибосомы) [ 33].Кроме того, ALKBH8, метилтрансфераза, участвующая в заключительной стадии образования mcm 5 s 2 U и mcm 5 U (Рисунок 1), высоко экспрессируется при раке мочевого пузыря, а нокдаун ALHBH8 вызывает гибель раковых клеток из-за снижения экспрессии. антиапоптотического белка сурвивина [29, 149]. Следовательно, многочисленные доказательства указывают на модификации U 34 , способствующие росту и метастазированию раковых клеток за счет обеспечения эффективного репрограммирования трансляции при переходе от нормального режима роста клеток к раку.В дополнение к повышенной регуляции генов Elongator, ALKBH8 и тиолаз CTU1 и CTU2, другие гены модификации тРНК, включая NSUN2 (m 5 C) и METTL1 (m 7 G), также становятся сверхэкспрессированными при раке человека [52,54,154,155] (Таблица 1). Более того, последние два гена участвуют в устойчивости к противораковой терапии [156], поскольку активация NSUN2 коррелирует с плохим прогнозом у пациентов с плоскоклеточной карциномой головы и шеи [157]. Изменения в модификации эпитранскриптомной тРНК в раковых клетках могут иметь большое значение для прогноза прогрессирования заболевания и лечения.Следовательно, более глубокое понимание основных путей, участвующих в модификации U 34 , необходимо для дальнейшего понимания его двустороннего характера при заболеваниях человека. Остается показать, можно ли идентифицировать и разработать специфический ингибитор элонгатора. Прежде всего, таргетная терапия против элонгатора должна определять терапевтическое окно, которое позволяет лечить раковые клетки и одновременно избегает негативного воздействия на нейрональные ткани.7. Фосфорегуляция элонгатора с участием Kti12 и Kti14 / Hrr25
Несколько исследований на дрожжах и других модельных организмах показали, что модификации тРНК, в том числе зависимые от элонгатора, могут изменяться в ответ на прогрессирование клеточного цикла и различные стрессы окружающей среды.Это указывает на то, что модификации тРНК подлежат регуляции, а не конститутивно формируются [158,159,160]. В случае комплекса Elongator из дрожжей были описаны несколько дополнительных белков, которые влияют на его активность модификации тРНК, а именно Kti11-Kti14 / Hrr25 и Sit4 (Рисунок 2). В соответствии с этим было показано, что изофермент казеинкиназы 1 (CK1) (Kti14 / Hrr25), протеинфосфатазы типа 2A (Sit4 · Sap185 и Sit4 · Sap190) и взаимодействующий элонгатор (Kti12) влияют на состояние фосфорилирования самой большой субъединицы каркаса элонгатора. Elp1 [98, 161, 162, 163].В принципе, динамическое фосфорилирование может влиять на каталитическую субъединицу Elp3 элонгатора, ее локализацию или ее способность взаимодействовать с субстратными тРНК [164,165,166]. Хотя неясно, какую именно роль играет фосфорилирование Elp1, оно было предложено как «включение / выключение» активности модификатора Elongator U 34 , например, в ответ на условия роста или клеточный стресс. Учитывая, что трансляция некоторых мРНК действительно зависит от собственно модификации U 34 и, следовательно, настраивается с помощью элонгатора [130, 167, 168], и что модификации тРНК, включая зависимые от элонгатора, действительно колеблются, это повышает вероятность того, что элонгатор является частью контроля трансляции. механизм, который функционирует как модификатор U 34 .Такая роль, которая отражает то, что фосфорилирование Elp1 казеинкиназой Kti14 / Hrr25 в значительной степени положительно для работы Elongator, согласуется с фенотипами потери функции, связанными с мутациями hrr25 / kti14 с мертвой киназой, абляционными заменами фосфозита Elp1 и специфическим ингибированием АТФ. аналоги аналоговочувствительного варианта киназы Hrr25 (Hrr25-I82G) [164,165]. Хотя Hrr25 оперирует многими клеточными функциями [169], что затрудняет получение четкого представления о сигналах фосфорилирования Elp1, его киназная активность необходима для полной функциональности рибосом и U 34 , содержащих тРНК, что демонстрирует его важность в трансляции мРНК и синтезе белка [164, 165, 170, 171, 172] .Поскольку динамическое фосфорилирование IKAP / Elp1 также наблюдалось в клетках меланомы человека [32] и изменялось в ответ на доступность инсулина, фармакологическое вмешательство в этот процесс может представлять привлекательный вариант для подавления функции элонгатора и, возможно, нарушения пролиферации раковых клеток. фосфорегуляция Elp1, по-видимому, является белком-партнером Elongator Kti12 (Figure 2). Хотя точная роль Kti12 плохо определена, дрожжевой белок и его растительный ортолог (DRL1 / ELO4) несут мотивы N-концевой P-петли, которые типичны для нуклеотидсвязывающих киназ и NTPases [173,174,175,176,177].В соответствии с функциональной ролью этого домена, усечение P-петли запускает дефекты, типичные для мутантов Elongator, включая потерю модификации U 34 [98]. Важно, что Kti12 взаимодействует с киназой Hrr25 зависимым от элонгатора способом и тем самым, по-видимому, поддерживает фосфорилирование Elp1 [98,100,102,178]. Это основано на данных, показывающих, что делеции гена KTI12 отменяют взаимодействие элонгатора с Hrr25, вызывают дефекты модификации U 34 , типичные для мутантов elp (и hrr25), и запускают образование гипофосфорилированных изоформ Elp1, подобных киназо-мертвым клеткам hrr25 [101, 161, 165]. .8. Регуляция элонгатора посредством Kti11 / Dph4 и Kti13
Два дополнительных фактора, участвующих в регуляции элонгатора, — это Kti11 (псевдоним Dph4) и Kti13 (псевдоним Ats1) [179] (Рисунок 2). Kti11 осаждает элонгаторные субъединицы (Elp1, Elp2, Elp3, Elp5) и связывается с Kti13 [108,180,181] в гетеродимере, который, как было показано, влияет не только на активность модификации элонгатора U 34 , но также имеет отношение к пути посттрансляционной модификации белка [108,180,184,182,183] . Комплекс Kti11 / Dph4 • Kti13 участвует в переносе электрона на Elp3 для радикальной SAM-зависимой модификации U 34 с помощью комплекса Elongator [182,183].Было обнаружено, что удаление Kti11 (т.е. отсутствие потока электронов) устраняет активность модификатора U 34 элонгатора, в то время как потеря Kti13 значительно снижает функцию модификации тРНК до ~ 20% от уровней дикого типа [78,180,181]. Kti11 / Dph4 участвует вместе с Dph2, Dph3, Dph5, Dph5, Dph6 и Dph7 в синтезе дифтамида, посттрансляционно модифицированного остатка гистидина (His699 в дрожжах; His715 у людей), обнаруженного в факторе удлинения трансляции 2 (EF2) [185,186]. EF2 является важным фактором трансляции, который опосредует транслокацию рибосомы на мРНК во время элонгации трансляции.Поразительно, что модифицированный дифтамидом EF2 является мишенью для дифтерийного токсина (DT), который инактивирует фактор трансляции через ADP (аденозиндифосфат) -рибозилирование и, таким образом, вызывает гибель отравленных клеток [187, 188, 189]. Таким образом, отсутствие Dph4 / Kti11 приводит к полной устойчивости к летальной АДФ-рибослязной активности DT [180,182,185,186,190]. Эти данные ясно указывают на патологическую роль модификации дифтамида в контроле роста и пролиферации клеток. Физиологическая значимость EF2, модифицированного дифтамидом, менее очевидна [191, 192].Однако недавние функциональные и структурные анализы показывают, что EF2, модифицированный дифтамидом, поддерживает поддержание рамки считывания и снижает рибосомные ошибки [190,193,194,195,196,197]. На основании анализа структурно-функциональных функций Kti11 / Dph4 несет в себе домен связывания металла (железо, цинк) [182,183,198], который имеет важное значение. как для модификации U 34 , несущей тРНК, посредством элонгатора, так и для синтеза дифтамида на EF2 [182,183]. Подобно бактериальным рубредоксинам, Kti11 / Dph4 является окислительно-восстановительным белком, способным переносить электроны к железо-серным кластерам основной субъединицы Elongator Elp3 [86,89] и радикальному ферменту SAM Dph2 • Dph3, который необходим для первого стадия биосинтеза дифтамида [108,180,186,199].Донг и др. (2014) реконструировали реакцию in vitro, в которой Dph4 / Kti11 был способен подавать электроны в кластер [4Fe – 4S] SAM фермента Dph2 • Dph3 для восстановительного расщепления SAM и образования 3-амино-3- карбоксипропильный радикал (ACP) впоследствии использовали для образования ACP-EF2, первого промежуточного соединения дифтамидного пути [200,201,202,203]. Более того, была идентифицирована Dph4 / Kti11 редуктаза Cbr1, которая необходима для рециклирования электронного носителя и передаточной функции Dph4 / Kti11 как в пути модификации U 34 , так и в пути модификации дифтамида [204].Образование стабильного гетеродимера Kti11 / Dph4 • Kti13 предполагает участие не только Kti11, но также и Kti13 в образовании дифтамида [181,182,183]. Действительно, было показано, что дрожжевой штамм с делецией гена KTI13 обеспечивает защиту от ингибирования роста DT (см. Выше) [182]. Таким образом, помимо Kti11 / Dph4 и Cbr1, Kti13 может быть еще одним фактором, действующим как в радикальных путях SAM для зависимой от элонгатора модификации тРНК, так и в синтезе дифтамида на EF2 [182,183]. В таблице 1 перечислены тяжелые патологии у людей, которые связаны с Зависимые от элонгатора и связанные с ними дефекты модификации тРНК.Точно так же дефекты генов, участвующих в пути синтеза дифтамида у высших эукариотических модельных организмов, могут быть связаны с различными заболеваниями и синдромами (таблица 3), такими как туморогенез [205,206,207,208,209,210,211], умственная отсталость, черепно-лицевые аномалии [212,213] и синдром Миллера-Дикера ( MDS) [214], а также обструкция дыхательных путей и аномалии внешних половых органов [215] у людей. У мышей в сочетании с дифтамидными дефектами наблюдались эмбриональная летальность, отек, полайдактилия, укорочение челюсти, расщелина неба, некроз, апоптоз и дефекты развития плаценты, а также недоразвитие нейронов [214, 216, 219, 220].Кроме того, сообщалось, что у китайских хомячков чувствительность к окислительному стрессу коррелирует с неспособностью модифицировать дифтамидом EF2 [218]. Более того, нарушение пролиферации кишечных стволовых клеток строго связано с нарушением функций Dph2 и Dph5 у Drosophila melanogaster [217]. Следовательно, дефекты как радикальных путей SAM для модификаций тРНК, так и EF2, которые, как было показано, влияют на точность и эффективность элонгации трансляции мРНК и вызывают рибосомные ошибки, потенциально связаны с множеством заболеваний (таблицы 1 и 3) от несколько эукариот.С гетеродимером Kti11 / Dph4 • Kti13 и Cbr1-редуктазой, квалифицируемыми как главные кандидаты для соединения и, возможно, перекрестного сшивания U 34 и модификаций дифтамида, появляется новая перспектива регуляции связанных с заболеванием радикальных ферментов-модификаторов SAM с помощью общего механизма это включает в себя управление потоком электронов к их центрам железо-сера (и от них). Потребуются дальнейшие исследования, чтобы выяснить, может ли лекарственное вмешательство в функционирование Kti11 / Dph4 • Kti13 и / или Cbr1 обеспечить новые схемы вмешательства против болезненных синдромов, связанных с динамическими изменениями эпитранскриптомных модификаций тРНК, типичных для дефектов в элонгаторе или связанном пути синтеза дифтамида. .(PDF) Форстен-Вильямс К., Чуа С.С., Ньюджент М.А. .. Кинетика связывания FGF-2 с гепарансульфат-протеогликанами и передача сигналов киназы MAP. Дж. Теор Биол 233: 483-499
Горовиц А., Саймонс М., 1998b. Фосфорилирование цитоплазматического
хвоста синдекана-4 регулирует активацию протеинкиназы Са. J. Biol.
Chem. 273, 25548–25551.
Горовиц, А., Ткаченко, Э., Саймонс, М., 2002. Рост фибробластов
-специфическая модуляция клеточного ответа с помощью синдекана-4.J.
Cell Biol. 157, 715–725.
Ибрахими, О.А., Чжан, Ф., Хрстка, С.С., Мохаммади, М., Линхардт,
,Р.Дж., 2004. Кинетическая модель для сборки комплекса передачи сигналов FGF, FGFR и протеогликана
. Биохимия 43,
4724–4730.
Извольский, К.И., Чжун, Л., Вэй, Л., Ю, К., Нуджент, М.А., Кардосо,
W.V., 2003a. Гепарансульфаты, экспрессируемые в дистальном отделе легкого, составляют
–, необходимые для связывания Fgf10 с эпителием и для ветвления дыхательных путей
–.Являюсь. J. Physiol. Легочная клетка. Мол. Physiol. 285,
L838 – L846.
Извольский, К.И., Шойхет, Д., Янг, Ю., Ю., К., Нугент, М.А.,
Кардосо, В.В., 2003b. Взаимодействия гепарансульфат-FGF10 во время морфогенеза легких
–. Dev. Биол. 258, 185–200.
Холоденко, Б.Н., Демин, О.В., Моерен, Г., Хук, Дж. Б., 1999.
Количественное определение краткосрочной передачи сигналов рецептором фактора роста эпидермиса
. J. Biol. Chem. 274, 30169–30181.
Лаеммли, У.К., 1970. Расщепление структурных белков при сборке
головки бактериофага Т 4. Природа 227,
680–685.
Лауффенбургер, Д.А., Линдерман, Дж. Дж., 1993. Рецепторы: модели для
привязки, трафика и сигнализации. Oxford University Press, New
York.
Lee, D., Oh, ES, Woods, A., Couchman, JR, Lee, W., 1998.
Структура раствора цитоплазматического домена синдекана-4 и его
взаимодействие с фосфатидилинозитол-4,5-бисфосфатом .J. Biol.
Chem. 273, 13022–13029.
Maher, P.A., 1993. Ингибирование тирозинкиназной активности рецептора фактора роста фибробластов
ингибитором метилтрансферазы
50-метилтиоаденозин. J. Biol. Chem. 268, 4244–4249.
Мали, М., Элениус, К., Миеттинен, Х.М., Ялканен, М., 1993. Ингибирование
стимулирования роста, индуцированного основным фактором роста фибробластов, за счет сверхэкспрессии
синдекана-1. J. Biol. Chem. 268, 24215–24222.
Мацуда, К., Маруяма, Х., Го, Ф., Клифф, Дж., Итакура, Дж.,
Мацумото, Ю., Ландер, А.Д., Корч, М., 2001. Глипикан-1 —
сверхэкспрессируется при раке груди человека и модулирует митогенные эффекты
множественных гепарин-связывающих факторов роста в клетках рака груди
. Cancer Res. 61, 5562–5569.
Мерри, К.Л., Галлахер, Дж. Т., 2002. Новые взгляды на биосинтез гепарансульфата
из исследования мутантных мышей. Biochem.Soc. Symp.
69, 47–57.
Miyaji, K., Tani, E., Nakano, A., Ikemoto, H., Kaba, K., 1995.
Ингибирование 50-метилтиоаденозином роста клеток и тирозин
активность киназы, стимулированная фактором роста фибробластов рецептор в
глиомах человека. J. Neurosurg. 83, 690–697.
Моерен, Г., Маркевич, Н., Демин, О., Кияткин, А., Горянин, И.,
Хук, Дж. Б., Холоденко, Б.Н., 2002. Температурная зависимость
сигнального пути рецептора эпидермального фактора роста. сеть может быть учтена кинетической моделью
.Биохимия 41, 306–320.
Monsonego-Ornan, E., Adar, R., Rom, E., Yayon, A., 2002. Убиквитилирование рецепторов FGF
: зависимость от активности тирозинкиназы
и роль в подавлении регуляции. FEBS Lett. 528, 83–89.
Murakami, M., Horowitz, A., Tang, S., Ware, JA, Simons, M., 2002.
Дельта протеинкиназы C (PKC) регулирует активность PKCalpha
в зависимости от Syndecan-4 . J. Biol. Chem. 277,
20367–20371.
Ньюджент, М.A., Edelman, E.R., 1992. Кинетика связывания основного фактора роста фибробластов
со своим рецептором и гепарансульфатом
протеогликан: механизм кооперативности. Биохимия 31,
8876–8883.
Ньюджент, М.А., Иоззо, Р.В., 2000. Фактор роста фибробластов-2. Int. J.
Biochem. Cell Biol. 32, 115–120.
О, Э.С., Каучман, Дж. Р., Вудс, А., 1997a. Фосфорилирование серина
цитоплазматического домена протеогликана синдекана-2. Arch. Biochem.
Biophys. 344, 67–74.
О, Э. С., Вудс, А., Каучман, Дж. Р., 1997b. Мультимеризация цитоплазматического домена
синдекана-4 необходима для его способности
активировать протеинкиназу С. J. Biol. Chem. 272, 11805–11811.
Oh, ES, Woods, A., Lim, ST, Theibert, AW, Couchman, JR,
1998. Синдекан-4 протеогликановый цитоплазматический домен и фосфат-
тидилинозитол-4,5-бисфосфат координированно регулируют протеинкиназу
C. активность.J. Biol. Chem. 273, 10624–10629.
Парк П.У., Рейзес О., Бернфилд М., 2000. Гепаран
сульфатных протеогликанов на клеточной поверхности: селективные регуляторы лиганд-рецептор
встреч. J. Biol. Chem. 275, 29923–29926.
Park, C.S., Schneider, I.C., Haugh, J.M., 2003. Кинетический анализ передачи сигналов
тромбоцитарного фактора роста / фосфоинозитид-3-киназы /
Akt в фибробластах. J. Biol. Chem. 278, 37064–37072.
Перримон, Н., Бернфилд, М., 2000. Особенности гепарансульфата
протеогликанов в процессах развития. Nature 404, 725–728.
Rapraeger, A., Krufka, A., Olwin, B., 1991. Потребность в сульфате гепаран
для bFGF-опосредованного роста фибробластов и дифференцировки миобластов
. Science 252, 1705–1708.
Razandi, M., Pedram, A., Merchenthaler, I., Greene, G.L., Levin,
E.R., 2004. Рецепторы эстрогена плазматической мембраны существуют, и
функционируют как димеры.Мол. Эндокринол. 1 июля (epub).
Schlessinger, J., 2002. Лиганд-индуцированная рецептор-опосредованная димеризация
и активация рецептора EGF. Cell 110, 669–672.
Schlessinger, J., Lax, I., Lemmon, M., 1995. Регулирование роста
Активация факторапротеогликанами: какова роль рецепторов с низким сродством
. Cell 83, 357–360.
Шлессингер Дж., Плотников А.Н., Ибрагими О.А., Елисеенкова А.В.,
Йе, Б.К., Яйон, А., Linhardt, R.J., Mohammadi, M., 2000.
Кристаллическая структура тройного комплекса FGF-FGFR-гепарин показывает
двойную роль гепарина в связывании и димеризации FGFR. Мол.
Cell 6, 743–750.
Schoeberl, B., Eichler-Jonsson, C., Gilles, E.D., Muller, G., 2002.
Вычислительное моделирование динамики каскада MAP-киназы
, активируемого поверхностными и интернализованными рецепторами EGF. Nat.
Biotechnol. 20, 370–375.
Шварцман, С.Ю., Муратов, К. Б., Лауффенбургер, Д. А., 2002.
Моделирование и вычислительный анализ опосредованной рецептором EGF коммуникации
клеток в оогенезе дрозофилы. Разработка 129,
2577–2589.
Сорокин, А., Мохаммади, М., Хуанг, Дж., Шлессингер, Дж., 1994.
Интернализация рецептора фактора роста фибробластов ингибируется точечной мутацией
в тирозине 766. J. Biol. Chem. 269, 17056–17061.
Сперинде Г.В., Ньюджент М.А., 2000.Механизмы внутриклеточного процессинга FGF-2
: кинетический анализ роли протеогликанов гепарансульфата
. Биохимия 39, 3788–3796.
Steinfeld, R., Van Den Berghe, H., David, G., 1996. Стимуляция занятости рецептора-1 фактора роста фибробластов
и передача сигналов клетками
поверхностно-ассоциированных синдеканов и глипикана. J. Cell. Биол. 133,
405–416.
Ткаченко, Э., Саймонс, М., 2002. Кластеризация индуцирует перераспределение
корового белка синдекана-4 в мембранные домены рафта.J. Biol.
Chem. 277, 19946–19951.
Ткаченко, Э., Лутгенс, Э., Стэн, Р.В., Саймонс, М., 2004. Фибробласт
Эндоцитоз фактора роста 2в эндотелиальных клетках протекает через
синдекан-4-зависимую активацию Rac1 и Cdc42-зависимую
макропиноцитарный путь. J. Cell Sci. 117, 3189–3199.
Volk, R., Schwartz, J.J., Li, J., Rosenberg, R.D., Simons, M., 1999.
Роль цитоплазматического домена синдекана в базовой передаче сигнала фибробластов
, зависимой от фактора роста.J. Biol. Chem. 274,
24417–24424.
Уорбертон, Д., Шварц, М., Теффт, Д., Флорес-Дельгадо, Г., Андер-
сын, К.Д., Кардосо, В.В., 2000. Молекулярная основа морфогенеза легких
. Мех. Dev. 92, 55–81.
СТАТЬЯ В ПРЕССЕ
K. Forsten-Williams et al. / Журнал теоретической биологии 233 (2005) 483–499498
Паклобутразол как регулятор роста растений | Химические и биологические технологии в сельском хозяйстве
Замедлители роста растений — это соединения, которые используются для замедления роста растений без изменения моделей развития и являются фитотоксичными [14].Самую большую группу замедлителей роста растений составляют химические вещества, антагонистические гиббереллинам (ГА), гормону, отвечающему за рост растений [7]. Коммерчески используемые ингибиторы биосинтеза GA: (а) соединения ониевого типа, (б) соединения с N -гетероциклом (триазольного типа), (в) структурные имитаторы 2-оксоглутаровой кислоты и (г) 16, 17-дигидроГА [14].
PBZ, входящий в группу триазольных регуляторов роста растений, широко используется в сельском хозяйстве [15]. Это ингибитор удлинения клеток и удлинения междоузлий, который замедляет рост растений за счет ингибирования биосинтеза гиббереллинов.Гиббереллины стимулируют удлинение клеток. Когда продукция гиббереллина подавляется, деление клеток все еще происходит, но новые клетки не удлиняются. В результате получаются побеги с одинаковым количеством листьев и междоузлий, сжатые до более короткой длины. Также наблюдалось уменьшение диаметра ствола и ветвей. Другой ответ деревьев на обработку PBZ — это увеличение выработки гормона абсцизовой кислоты и фитола, компонента хлорофилла, которые полезны для роста и здоровья деревьев.PBZ может также вызывать морфологические модификации листьев, такие как более мелкие устьичные поры, более толстые листья и увеличенное количество и размер поверхностных придатков, а также повышенная плотность корней, что может обеспечить улучшенную устойчивость к стрессу окружающей среды и устойчивость к болезням [16]. PBZ также обладает некоторой фунгицидной активностью из-за его способности как триазола ингибировать биосинтез стеролов [16].
Химия
PBZ ([(2R, 3R + 2S, 3S) -1- (4-хлорфенил) 4,4-диметил-2- (1,2,4-триазол-1-ил) пентан -3-ol]) был разработан как регулятор роста растений и зарегистрирован под торговыми названиями, такими как Bonzi, Clipper, Cultar и Parsley.Он относится к триазольным соединениям, которые характеризуются кольцевой структурой, содержащей три атома азота, хлорфенил и боковые углеродные цепи [17]. Структурно PBZ представляет собой замещенный триазол с двумя асимметричными атомами углерода и образуется в виде смеси энантиомеров 2R, 3R и 2R, 3R и 2S, 3S [18] (рис. 1).
Рис. 1Структура паклобутразола
Механизм действия
Хотя точные особенности молекулярной структуры, которые обеспечивают регуляторную активность роста растений, не совсем понятны, похоже, что это связано со стереохимическим расположением заместителей на углеродная цепь [19].Есть указания на то, что энантиомеры, имеющие S-конфигурацию у хирального углерода, несущего гидроксильную группу, являются ингибиторами биосинтеза GA. Один из ингибиторов биосинтеза ГА, паклобутразол, в основном используется в качестве замедлителя роста и защиты от стресса [20]. Это замедление роста связано с вмешательством PBZ в биосинтез гиббереллина путем ингибирования окисления энт-каурена до энт-кауроновой кислоты посредством инактивации цитохром Р450-зависимой оксигеназы [21, 22]. Кроме того, он имеет тенденцию быть намного более эффективным, чем другие регуляторы роста растений, при относительно низкой скорости применения [22].
PBZ также влияет на синтез гормона абсцизовой кислоты и фитола. Абсцизовая кислота также синтезируется терпеноидным путем (рис. 2). Когда синтез гиббереллинов блокируется, больше предшественников терпеноидного пути накапливаются и шунтируются, способствуя генезу абсцизовой кислоты [23]. Сообщалось также, что он ингибирует нормальный катаболизм АБК [24]. Воздействие ПБЗ на процессы синтеза и катаболизма приводит к повышению концентрации АБК в листьях.Одна из основных ролей АБК — закрывать устьичную апертуру и уменьшать потерю воды листьями из-за транспирации. Улучшение водных отношений в обработанных растениях происходит из-за увеличения содержания АБК, которое уменьшает устьичную апертуру, уменьшает рост побегов и приводит к уменьшению площади поверхности для транспирации, большему количеству корней для поглощения воды и анатомическим изменениям в листьях, которые создают препятствия для потери воды.
Рис. 2Терпеноидный путь биосинтеза гиббереллинов, абсцизовой кислоты, фитола и стероидов, а также путь деградации абсцизовой кислоты.Шаги, блокируемые паклобутразолом, обозначены геранилдифосфатсинтазой (GPS), фарнезилдифосфатсинтазой (FPS), геранилгеранилдифосфатсинтазой (GGPS), энт-копалилдифосфатсинтазой (CPS), энт-кауренсинтазой (Kaurene-оксидом) (KO), оксидаза энт-кауреновой кислоты (KAO), геранилгеранилредуктаза (GGRS), хлорофилл-синтаза (CHL) и фитоен-синтаза (PSY) являются ферментами, участвующими в пути терпеноидов. ABA 8′-гидроксилаза (ABA 8’OH) участвует в ферментативном расщеплении ABA до фазовой кислоты.KO, KAO и ABA 8’OH — ферменты, ингибируемые при применении PBZ.
Транслокация и химическая стабильность
Ранее считалось, что триазолы в основном транспортируются акропетально в ксилеме [25]. Однако PBZ был обнаружен в ксилеме и соке флоэмы клещевины [26] и груши [27], что указывает на то, что триазолы могут переноситься акропетально и базипетально. Хотя метаболическая судьба применяемых препаратов не исследована подробно, большинство из них обладают высокой химической стабильностью [28] и, в зависимости от места применения, имеют тенденцию к медленному метаболизму [15].Ранний и Мартин [29] наблюдали более быстрый метаболизм PBZ в листьях яблони, чем в других частях растения, в то время как Стерретт [30] обнаружил мало доказательств метаболизма PBZ в проростках яблони. PBZ сравнительно более устойчив к деградации, чем BAS 111 [31].
Способы нанесения
Наиболее распространенными методами нанесения PBZ являются опрыскивание листвой и замачивание средой. PBZ показывает хорошие результаты для обоих методов [32]; однако поливы действуют дольше и обеспечивают равномерный контроль высоты растений при более низких дозах [33].Когда PBZ применяется путем опрыскивания листвы, соединение плохо растворяется в воде и, следовательно, мало перемещается во флоэме. Таким образом, при нанесении распылением на полог растения его действие ограничивается областью влажного контакта [34]. С другой стороны, нанесение PBZ смачиванием является равномерным и увеличивает эффективность продукта при более низких концентрациях по сравнению с опрыскиванием листьев. Более того, обливание PBZ может напрямую ингибировать синтез GA, поскольку корни синтезируют большие количества GA [35].Аналогичным образом Banon et al. [36] и AlKhassawneh et al. [37] продемонстрировали, что обводнение было более эффективным, позволяя использовать меньшее количество PBZ, что желательно как по экологическим, так и по экономическим причинам. Такая эффективность может быть напрямую связана с его высокой стойкостью в орошении почвы [38] и в органах растений [39, 40]. Гент и МакЭвой [41] также указали, что PBZ сохраняется в однолетних, травянистых многолетних и особенно древесных декоративных растениях.
PBZ считается неподвижным химическим веществом флоэмы [7], хотя существуют некоторые прямые [26] и косвенные [42] доказательства того, что он частично подвижен во флоэме.Исследования показывают, что PBZ и униконазол-P перемещаются в растениях акропетально через ксилему, накапливаются в листьях и имеют очень низкую подвижность во флоэме [14]. Это приводит к низкому уровню остатков PBZ в семенах и плодах, поскольку они снабжаются питательными веществами через флоэму [25]. Однако низкая подвижность PBZ во флоэме дополнительно снижает эффективность опрыскивания листвы, поскольку действие PBZ на рост растений будет ограничиваться местом нанесения.
Нормы внесения
Многое было сделано для определения наилучшей нормы внесения PBZ в разных местах.При определении нормы внесения PBZ следует учитывать такие факторы, как возраст деревьев, степень вегетативного роста и метод внесения. Ставки также по-разному влияют на различные параметры дерева. В целом количество PBZ, необходимое для стимулирования цветения и плодоношения плодовых культур, очень низкое [27].
Норма внесения в почву зависит от размера дерева и сорта. Норма определяется путем умножения диаметра кроны дерева в метрах на 1–1,5 г активных ингредиентов PBZ [43].Они указали, что другие факторы, включая тип почвы, ирригационную систему и т. Д., Могут влиять на активность PBZ и, таким образом, могут быть необходимы для повышения эффективности химического вещества. Что касается них, передозировка может вызвать нежелательные эффекты, такие как ограниченный рост, деформация метелки (слишком компактная) и деформация побегов. Они также утверждали, что для обеспечения равномерного цветения и уменьшения вредных побочных эффектов был исследован поиск лучших методов нанесения, и один из подходов заключается в применении большого объема с низкой концентрацией PBZ для улучшения лучшего покрытия.
Оптимизация дозы PBZ является предпосылкой для любых программ повышения урожайности. Серьезная и нежелательная потеря семян и выхода масла Camelina наблюдалась, когда растения обрабатывали более высокой концентрацией PBZ (125 мг L -1 ), в то время как доза PBZ составляла от 75 мг L -1 до 100 мг L . −1 может эффективно улучшить экономические характеристики, в том числе более высокие урожаи семян и масла у Camelina . Сильное замедление роста Camelina также отразилось на высоте растений, ветвях и размере кроны при опрыскивании растений более высокой концентрацией PBZ (125 мг л –1 ) [44].Он также сообщил, что урожай семян Camelina увеличился на 74,23% по сравнению с контролем с внесением 100 г L -1 . Точно так же снижение урожайности было зарегистрировано для арахиса [45] и ятрофы [46], связанного с более высокими концентрациями PBZ.
Камран и др. [47] описали, что замачивание семян под 300 мг L -1 PBZ увеличивало средний урожай зерна кукурузы на 61,3% по сравнению с контролем. Патил и Талати [48] также сообщили, что внесение 5 г PBZ в почву позволило вызвать раннее и регулярное плодоношение с 2.Увеличение урожайности манго вар. В 8 раз. Альфонсо. Кроме того, PBZ в количестве 150 мг L -1 в бутылочной тыкве, 100 мг L -1 в горькой тыкве, 150 мг L -1 в французской фасоли, 125 мг L -1 в огурце. и 40 мг L -1 в томате повысили урожайность и качество плодов [49].
Ответ растений на PBZ
Биосинтез растительных гормонов
Гиббереллин: Гиббереллины (ГА) представляют собой большое семейство тетрациклических дитерпеноидных регуляторов роста растений.С момента первоначального открытия> 130 ГА было идентифицировано в растениях, грибах и бактериях, хотя только несколько ГА обладают биологической активностью [50]; Многие небиоактивные ГА существуют в растениях, и они действуют как предшественники биоактивных форм или деактивированные метаболиты. Гиббереллины (ГА) представляют собой гормоны растений, которые необходимы для многих процессов развития растений, включая прорастание семян, удлинение стебля, разрастание листьев, развитие трихом, созревание пыльцы и индукцию цветения [51].Основные биоактивные ГА, которые включают GA 1 , GA 3 , GA 4 и GA 7 , происходят из основного каркаса дитерпеноид карбоновой кислоты и обычно имеют гидроксильную группу C 3 [50] .
Соединения триазола антагонистичны гиббереллинам и ауксинам, уменьшая удлинение и деление клеток за счет ингибирования биосинтеза GA 3 [52]. Они проявляют разную степень роста растений и фунгицидной активности. Интенсивность их биологической активности зависит от их изомерной формы [17].Свойство PBZ задерживать рост в значительной степени объясняется вмешательством в биосинтез гиббереллинов. Гиббереллины синтезируются из мевалоновой кислоты через изопреноидный путь, а PBZ специфически ингибирует окисление энт-каурена до энт-кауреновой кислоты посредством инактивации цитохром Р-450-зависимых оксигеназ [18]. Кроме того, ингибирование роста, индуцированное PBZ, может быть отменено экзогенным применением гиббереллинов [53]. Эти наблюдения подтверждают гипотезу о том, что ингибирование роста из-за PBZ в первую очередь связано со сниженным биосинтезом гиббереллинов.
Абсцизовая кислота: влияние PBZ на ABA представляет интерес, потому что ABA, как и гиббереллины, синтезируется через изопреноидный путь, и эти два соединения часто проявляют противоположные физиологические активности. Действие PBZ на ABA может быть источником защиты от стресса, которая наблюдалась с PBZ [19]. ABA — это естественный регулятор роста растений, который участвует в акклиматизации растений и защите от стресса окружающей среды. Было показано, что экзогенное применение АБК повышает устойчивость растений к засолению, озону, жаре, холоду и замораживанию [54].
Mackay et al. [54] продемонстрировали, что PBZ индуцировал стрессоустойчивость, а также увеличивал эндогенные концентрации ABA в фасоли. Hauser et al. [55] также продемонстрировали, что PBZ значительно увеличивает уровни эндогенной АБК в отслоившихся листьях и проростках масличного рапса, выращенных на гидропонике. АБК накапливалась пропорционально концентрации PBZ. Mackay et al. [54] также предположили, что защита от стресса, обеспечиваемая PBZ, может частично быть результатом их влияния на эндогенные концентрации ABA.Однако оба эксперимента показали, что повышение АБК было кратковременным и в конечном итоге снизилось до нормального или ниже контрольного уровня. Hauser et al. [55] предположили, что это может быть связано со стимулированием катаболизма АБК и / или ингибированием ее биосинтеза. Следовательно, обеспечение непрерывного снабжения PBZ в течение вегетационного периода может помочь поддерживать более высокие уровни эндогенной АБК и тем самым продлить ее защитное действие от стресса. Кроме того, Али и Латиф [56] также сообщили, что PBZ увеличивает эндогенный уровень АБК в пшенице.
Цитокинин: Цитокинины синтезируются в корнях и акропетально перемещаются к побегам, где они регулируют как развитие растений, так и старение [57]. Они участвуют в контроле различных процессов развития растений, таких как деление клеток, апикальное доминирование, устьичное поведение, формирование корней, старение листьев и развитие хлоропластов [58].
Zhu et al. [21] наблюдали повышение уровня эндогенного цитокинина (зеатина) в ксилемном соке молодых деревьев яблони в ответ на обработку PBZ.Обработка PBZ задерживала начало старения винограда [59] и черники [60]. Сообщалось, что цитокинин или химические вещества, подобные тидиазурону, с цитокининоподобной активностью, стимулируют синтез хлорофилла и замедляют старение [61], и, таким образом, физиологические реакции, индуцированные PBZ, могут быть связаны с повышенным синтезом цитокинина или предотвращением его деградации.
Fletcher et al. [7] также предположили, что триазолы стимулируют синтез цитокининов и усиливают дифференциацию хлоропластов, биосинтез хлорофилла и предотвращают деградацию хлорофилла.Сообщалось о повышенном уровне цитокининов и полиаминов по сравнению с гормонами, способствующими старению, ABA и этиленом, у растений, обработанных PBZ. PBZ замедляет старение и увеличивает период «зеленого цвета» у Camelina sativa [44] за счет повышения эндогенных уровней цитокининов, что способствует образованию хлорофилла и повышению активности некоторых антиоксидантных ферментов. Более длительный «зеленоватый» характер одновременно увеличивал период фотосинтеза листьев у растений, применявших PBZ, за счет сохранения фотосинтетической эффективности листьев в течение более длительного времени, что, в свою очередь, увеличивало продуктивность растения Camelina [44].
Защита от стресса
Биохимические эффекты триазола включают детоксикацию активных форм кислорода, повышенное содержание антиоксидантов и хлорофилла (Chl) [22]. Совсем недавно было обнаружено, что соединения триазола защищают растения от различных стрессов окружающей среды, включая переохлаждение, засуху, жару, заболачивание, загрязнители воздуха и тяжелые металлы [7, 62]. Опосредованная триазолом защита от стресса часто объясняется гормональными изменениями, такими как повышение цитокининов, кратковременное повышение АБК и снижение этилена [19, 54].Повышенная толерантность к холоду в обработанных триазолом помидорах [64] была связана с повышением концентрации антиоксидантных ферментов. В обработанных помидорах, помимо увеличения антиоксидантов альфа-токоферола и аскорбата, количество свободных жирных кислот было выше, и наблюдалось снижение потери мембранных фосфолипидов по сравнению с необработанными контролями. PBZ предотвращает снижение общего содержания хлорофилла в растениях кукурузы после воздействия низких температур [64]. PBZ-индуцированная толерантность к низкотемпературному стрессу была связана с повышенным уровнем эндогенной АБК [7], которая, как сообщается, запускает генетические процессы закаливания.В полевых исследованиях ПБЗ повысил зимостойкость гороха и зерновых культур [25] и устойчивость кукурузы к морозам.
PBZ увеличивает выживаемость растений в условиях засухи за счет ряда физиологических реакций. Снижение скорости транспирации (из-за уменьшения площади листьев), повышенное диффузионное сопротивление, уменьшение водного потенциала, повышенное относительное содержание воды, меньшее потребление воды и повышенная антиоксидантная активность — вот некоторые из известных ответов [21, 65].PBZ также защищает растения от повреждений, вызванных высокой температурой [64, 65]. Защита от высокотемпературного стресса сопровождается выработкой стрессовых белков с низкой молекулярной массой [66] и повышением активности антиоксидантных ферментов [64].
Также сообщалось, что некоторые факторы окружающей среды, такие как засуха, низкая и высокая температура, могут вызывать избыток токсичных свободных от кислорода радикалов [67]. Сообщается, что некоторые из ферментов, улавливающих свободные радикалы, увеличиваются в растениях пшеницы [65] и кукурузы [64] после обработки PBZ, и их активность сохраняется даже после воздействия экстремальных температур.Соединения триазола увеличивают способность обработанных растений улавливать свободные радикалы, включая уровни каротиноидов, аскорбата, супероксиддисмутазы и аскорбатпероксидазы [63]. Берова и др. [68] предположили, что защита, вызванная PBZ, была вызвана аналогичным механизмом усиленных систем улавливания свободных радикалов.
Разделение ассимилятов
Разделение ассимилятов по разным стокам может контролироваться экологически регулируемым гормональным балансом [69]. Обработка PBZ увеличивала соотношение корней к побегам [64], увеличивала разделение ассимилятов на экономически важные части растения, такие как луковицы [70, 71], клубни картофеля [72, 73], корень моркови [8] и урожайность зерна риса [ 74].Механизм клубней действовать как доминирующий поглотитель во время разделения ассимилятов может быть связан со стимулированным PBZ низким уровнем GA в ткани клубня, который увеличивает активность поглощения клубней [72].
Setia et al. [75] также сообщили, что применение PBZ привело к общему увеличению сухой массы растения и лучшему распределению ассимилятов (процентное соотношение сухой массы кремнезема к сухому веществу растения) в Brassica juncea и Brassica carinata . Аналогичным образом Kumar et al.[45] сообщили, что обработка PBZ увеличивала урожай семян у Camelina sativa , и это повышение урожайности коррелировало с улучшением физиологии ассимиляции CO 2 , распределения активности поглощения ассимилятов и укоренения.
Кроме того, Yeshitela et al. [76] сообщили, что более высокие уровни PBZ подавляли вегетативный рост манго, а ассимилят, который должен был быть потрачен на вегетативный рост, был направлен на усиление цветения. Об этом свидетельствует более высокий уровень общих неструктурных углеводов в побегах обработанных деревьев перед цветением.Точно так же снижение вегетативного роста винограда за счет изменения относительной силы поглощения внутри растения имело косвенное последствие, позволившее большему распределению ассимилятов для репродуктивного роста, образования цветочных бутонов, образования плодов и роста плодов обработанных растений [77].
Поглощение минералов
Влияя на морфологию побегов и корней, PBZ изменяет поглощение минералов. Ригер [78], работавший в гидропонике на подвоях персика Nemaguard, обнаружил, что обработка PBZ вызывает снижение N, P, K, Fe и Mo, тогда как уровни Ca, Mg, B и Mn повышаются за счет PBZ.Этот автор утверждал, что величина изменений в подкормке листьев пропорциональна степени подавления роста. В случае яблонь Фудзи Хуанг и др. [79] обнаружили, что разница в общем количестве сухого вещества, накопленного на 1 кг листьев, незначительна. С другой стороны, Wang et al. [80] наблюдали, что обработка PBZ увеличивает содержание N, P, K, Ca, Mg, B и Zn в листьях груши. Ригер и Скалабрелли [81] продемонстрировали на персиковом дереве, что концентрации N, P, K и Fe в листве немного снижаются, а концентрации Ca, Mg, B и Mn увеличиваются.Недавно Yeshitela et al. [76] также сообщили, что PBZ увеличивает содержание Mg, Cu, Zn и Fe в листьях манго, не влияя на концентрацию N, P, K и Ca. Кроме того, этот автор указал, что более высокая концентрация PBZ (8,25 г действующего вещества / дерево) приводила к снижению концентрации Cu, в то время как увеличение концентрации PBZ (2,75-8,25 г действующего вещества / дерево) не показывало увеличения концентрации Zn.
Рост, урожайность и качество растений
Прорастание и развитие проростков
Общие проблемы, обнаруживаемые при обработке семян регуляторами роста, — это снижение или отсутствие прорастания и задержка появления всходов.Семена яблони ( Pyrus malus Mill.), Пропитанные 7 мг раствора PBZ -1 , имели 35% ингибирование прорастания и задержку прорастания на 2 дня [82]. Аналогичным образом семена миндаля ( Prunus dulcis L.), пропитанные растворами PBZ на 4000 или 8000 мг / л –1 в течение 15 мин, не прорастали [83].
Процент прорастания семян томатов, замачиваемых в 500 или 1000 мг L −1 PBZ в течение 6, 16 или 24 часов, был ниже, чем у замаченных в воде семян [84]. Кроме того, они обнаружили, что снижение высоты проростков через 36 дней после посева было> 30% для семян, которые были замачены в течение 16 часов в 500 или 1000 мг L -1 PBZ по сравнению с теми, которые были замачены в воде.Пилл и Гюнтер [85] также обнаружили, что воздействие на семена Cosmos bipinnatus 1000 мг / л −1 PBZ во время замачивания или замачивания снижает высоту проростков, а также снижает всхожесть проростков, причем реакции больше при более длительном воздействии во время замачивания, чем во время замачивания. Точно так же Пазиан и Беннетт [84] отметили, что 500 или 1000 мг L -1 PBZ снижали и замедляли прорастание семян томатов, герани и календулы. Это может быть связано с PBZ, который прилипает к семенной оболочке обработанных семян, а затем диффундирует в питательную среду, где он может поглощаться корнями проростков [86].Однако PBZ может проникать через семенную оболочку и оказывать прямое токсическое действие на зародыш. Покровы семян томатов имеют полупроницаемый слой [87], который может препятствовать проникновению PBZ в эндосперм и зародыш, тем самым уменьшая негативное влияние PBZ на прорастание.
Кроме того, Кар и Гупта [88] описали, что обработка семян подсолнечника ( Helianthus annus L.) и сафлора ( Carthamus tinctorius L.) PBZ снижает скорость прорастания и замедляет рост проростков.Аналогичным образом обработка семян ПБЗ (250 мг на 1 кг семян) замедляла удлинение первичных листьев у пшеницы ( Triticum durum L.), ячменя ( Hordeum vulgare L.), овса ( Avena sativa L.) , рожь ( Secale cereale L.) [89]. Первые настоящие листья проростков из семян, обработанных триазолом, имели нарушенную морфологию. У этих сеянцев удлинение корней было менее сильно замедлено, чем рост побегов; корни были толще и содержали больше воды [89].
Для семян сельдерея, которым необходим свет для прорастания, действие PBZ на биосинтез GA также зависело от света.PBZ оказывал слабое влияние на прорастание семян на свету, что могло быть связано с увеличением чувствительности семян к GA и, соответственно, более низкой потребностью в GA для прорастания [90]. Другие данные также предполагают, что прорастание семян, обработанных PBZ, зависит от уровней GA в семенах, на которые влияет интенсивность света [91].
Добавление GA или других химикатов (этифона) к семенам может преодолеть влияние регуляторов роста на прорастание семян. Только 5% семян петунии ( Petunia hybrida L.) прорастали после их впитывания в течение 14 дней в агаризованной среде, содержащей 5 мг L -1 PBZ.Добавление 10 мкМ раствора GA к субстрату, обработанному PBZ, улучшило всхожесть петунии до 65% [92]. Вымачивание в растворах PBZ ингибировало прорастание семян амаранта ( Amaranthus sp.), В то время как дальнейшее замачивание семян в гиббереллине, этифоне или 1-аминоциклопропан-1-карбоновой кислоте обращало ингибирующее действие PBZ [93].
Рост побегов
Обработка триазолом обычно уменьшает длину побегов и увеличивает толщину стебля молодого растения, а также ускоренное формирование корней является значительным преимуществом обработки паклобутразолом в Lycopersicon esculentum [94].Обработка триазолом оказывала более выраженный эффект уменьшения высоты растений пшеницы и выглядела более зеленой [6].
Самая яркая реакция роста, наблюдаемая у разных видов, обработанных PBZ, — это снижение роста побегов [95]. Этот отклик можно объяснить, прежде всего, уменьшением длины междоузлия. Было также обнаружено, что PBZ эффективно ингибирует высоту растений, разрастание листьев и изменение стебля у Syzygium campanulatum [96]. Точно так же высота растений была значительно уменьшена при применении PBZ у канолы [97], Vigna radiata [98], Epidendrum radicans [40], манго [99], пшеницы [68], Dianthus caryophyllus [36], Sesamum indicum [100] и Ocimum sanctum [101].
PBZ эффективно подавляет рост широкого диапазона видов растений, при этом обработанные растения, как правило, меньше и компактнее на вид и имеют более темные зеленые листья [102,103,104,105]. Терри и Милли [106] и Себастьян и др. [107] также сообщили, что растения, обработанные PBZ, имеют тенденцию быть темно-зелеными, более короткими и более компактными на вид. Точно так же обработка растений хризантемы PBZ в качестве увлажнения почвы приводила к получению более толстых листьев, уменьшенного диаметра стебля и увеличения диаметра корней [108].Модификация роста побегов с помощью PBZ может быть полезной для максимизации отдачи на единицу земли, позволяя увеличить популяции компактных растений на единицу площади земли.
Рост листа
Площадь листа: Обработка триазолом значительно уменьшила площадь листа, и это уменьшение могло быть связано с уменьшением размера листа [109]. Gomathinayagam et al. [110] также сообщили, что обработка триазолом уменьшала площадь листьев у Manihot esculenta .
PBZ вызывает различные морфологические модификации в зависимости от вида растений, стадии роста, скорости и способа применения [76, 107].Виджаялакшим и Сринивасан [111] обнаружили, что применение PBZ в манго оказалось значительно лучше в увеличении площади листьев по сравнению с другими обработками, такими как нитрат калия, мочевина и этрел, при средней площади 94,89 см 2 , тогда как контроль был только 63,65 см 2 . По мнению этих авторов, увеличение площади листьев позволило преодолеть ограничение истощения запасов пищевых материалов. Поскольку запасных кормовых материалов тогда было в большом количестве, удалось добиться прерывания чередующегося цикла плодоношения у выбранных сортов.Однако было обнаружено, что это противоречит выводам Fernandez et al. [112], которые сообщили об уменьшении площади листьев с помощью PBZ у Phillyrea angustifolia . Точно так же обработка паклобутразолом также уменьшила площадь листьев у Solanum tuberosum [72], Ocimum sanctum [101], Hordeum vulgare [113], Catharanthus roseus [114] и растений циннии [115]. Хотя PBZ уменьшил площадь поверхности растений, он улучшил жизнеспособность листьев; Таким образом, уменьшение площади поверхности листьев было компенсировано отсутствием опадания листа и его долговечностью [72].
Синтез хлорофилла Несколько исследований продемонстрировали увеличение содержания хлорофилла в растениях, обработанных триазолом [7, 94]. Эффект озеленения, вызванный обработкой растений регуляторами роста, можно объяснить увеличением содержания хлорофилла и / или более плотной упаковки хлоропластов на единицу площади листа из-за уменьшения площади листа [116]. Аналогичное объяснение предлагается повышенному содержанию хлорофиллов a и b в листьях картофеля [72].
Dewi et al. [117] сообщили, что растения черного риса, обработанные 25 или 50 ppm PBZ, имеют более зеленые листья по сравнению с контролем, и листья также испытали позднее старение.Это может быть связано с увеличением активности окислительных ферментов, препятствующих созреванию клеток. Аналогичным образом, исследования ятрофы [46], tef [118] и Camelina [119] показали, что хлорофилл был выше на растениях, обработанных PBZ, по сравнению с контролем. Повышенное содержание хлорофилла, обработанного PBZ, может быть связано с минимизацией повреждений, вызванных реактивным кислородом и изменениями уровней каротиноидов, аскорбата и аскорбатпероксидазы.
Отчет Nivedithadevi et al.[120] показали, что растения, обработанные PBZ, синтезировали больше цитокинина, что, в свою очередь, усиливало дифференциацию хлоропластов и биосинтез хлорофилла, а также предотвращало деградацию хлорофилла. Берова и Златев [94] также сообщили, что увеличение содержания хлорофилла может быть приписано более высокому содержанию цитокинина, который, как известно, стимулирует биосинтез хлорофилла и / или снижает катаболизм хлорофилла. Более того, PBZ, по-видимому, задерживает начало старения, которое выражается скоростью деградации хлорофилла в прикрепленных листьях маша, что, вероятно, связано с повышенным эндогенным уровнем цитокининов из-за их вторичного воздействия на растения [7].У некоторых видов растений листья, обработанные PBZ, сохранялись дольше, а начало старения значительно замедлялось [59, 60]. Активность, задерживающая старение, может быть связана с влиянием PBZ на содержание эндогенных цитокининов [7].
Скорость фотосинтеза и скорость транспирации Были опубликованы противоречивые отчеты о влиянии PBZ на эффективность фотосинтеза сельскохозяйственных культур; однако косвенно, уменьшая площадь листьев, это может уменьшить площадь фотосинтетической поверхности и, таким образом, уменьшить фотосинтез всего растения [25].ДеДжонг и Дойл [121] отметили отсутствие явного влияния PBZ на скорость фотосинтеза хорошо открытых нектариновых листьев в течение большей части вегетационного периода. Davis et al. [25] сообщили, что скорость фотосинтеза снизилась в результате ингибирования роста листьев. Лист — важная часть растения, поскольку он содержит клетки мезофилла, которые специализируются на фотосинтезирующих тканях. Gaussoin et al. [122] также сообщили, что растения, обработанные замедлителями роста растений, часто оказывали умеренное сдерживающее действие на скорость обмена углекислого газа, таким образом, возможно, снижая скорость фотосинтеза.
Ахмад Назарудин и др. [123] описали, что скорость фотосинтеза и скорость транспирации Syzygium myrtifolium были снижены после обработки PBZ. Это может быть связано со снижением скорости фотосинтеза, что также будет означать снижение скорости транспирации, поскольку оба процесса связаны с открытием и закрытием устьиц [124]. Снижение скорости транспирации уменьшило бы процент воды, выделяемой через устьица. Олсен и Андерсен [125] сообщили, что снижение скорости транспирации защитит растение от абиотического стресса из-за ограничения воды или периода засухи.Кроме того, PBZ повышал устойчивость растений к стрессу за счет увеличения потенциала давления ксилемы обработанных растений и, таким образом, повышения уровня влажности растений в период засухи. Согласно Abod и Jeng [126], на снижение скорости фотосинтеза и скорости транспирации влияли активность устьиц и площадь листьев. Однако Ахмад Назарудин и др. [123] сообщили, что активность устьиц не была основным фактором снижения скорости фотосинтеза и транспирации у Syzygium myrtifolium. Возможной причиной могло быть уменьшение площади листьев, что в дальнейшем способствовало уменьшению общей поверхности листьев, поглощающей солнечный свет.
Напротив, PBZ увеличивал скорость чистого фотосинтеза листа [72]. Это может быть связано с более высоким содержанием хлорофилла и более ранним клубнеобразованием в ответ на обработку PBZ. Об увеличении нетто-фотосинтеза в ответ на PBZ также сообщалось у Setaria italic [127] и конского каштана [128].Точно так же Сумья [129] сообщил, что применение PBZ для листвы и смачивания нута поддерживает более высокую скорость фотосинтеза в условиях водного дефицита и более быстрое восстановление после прекращения водного стресса. Увеличение межклеточной концентрации CO 2 и изменение устьичной проводимости были оценены как причины более высокого фотосинтеза у обработанных PBZ Amorphophallus campanulatus [20, 130] и красного огненного шипа в горшке [131].
Толщина листа: более высокое отложение эпикутикулярного воска на обработанных листьях может быть связано с увеличением эндогенных уровней АБК в ответ на обработку PBZ [23].Повышение уровня АБК стимулирует синтез белков-переносчиков липидов в ячмене, которые играют важную роль в образовании эпикутикулярных восков, процесс, который влияет на водное соотношение листьев [132]. Обработка PBZ вызвала увеличение общего количества парафина на 10% и изменила долю определенных парафинов в горшечных сортах роз в течение 11 дней после применения [133]. Развитие более толстого эпикутикулярного воскового слоя обеспечивает лучшую защиту от некоторых патогенов растений и незначительных механических повреждений [134].
Tekalign и Hammes [72] обнаружили, что растения картофеля cv. Земен лечился ПБЗ в дозе 67,5 мг д.в. на одно растение увеличилась общая толщина листьев с 215 до 267 мкм, что примерно на 24% больше, чем в контроле. Эти авторы также указали, что увеличение толщины листа объясняется увеличением диаметра эпидермальных клеток, длины палисадных клеток и глубины губчатого мезофилла. Sopher et al. [35] также сообщили, что у кукурузы на листьях, обработанных PBZ, наблюдалось больше эпикутикулярных отложений воска, они были толще и шире из-за увеличения сосудистых элементов, эпидермиса, мезофилла и клеток оболочки пучка.
В арахисе ( Arachis hypogaea L.) листья, обработанные PBZ, демонстрировали хорошо дифференцированные слои палисада и губчатого мезофилла с более длинными клетками [135]. Аналогичным образом Кишорекумар и др. [136] показали, что листья китайского картофеля ( Solenostemon rotundifolius ), обработанные PBZ, показали увеличенную толщину верхнего и нижнего эпидермиса, а также длину палисадных и губчатых клеток. Кроме того, Jaleel et al. [137] сообщили, что количество клеток в ткани палисада (мезофилла) на единицу площади увеличивалось в большей степени у обработанных PBZ Catharanthus roseus .
Рост стебля
Подавление высоты растения PBZ происходит потому, что соединение блокирует три отдельных этапа терпеноидного пути продукции гиббереллинов (GA). GA увеличивает удлинение междоузлий интактных стеблей [124]. Liu и Loy [138] показали, что GA способствует делению клеток, стимулируя клетки в фазе G 1 для перехода в фазу S и сокращая продолжительность фазы S. Они пришли к выводу, что увеличение количества клеток приводит к более быстрому росту ствола. Но обработка растений PBZ приводила к получению стеблей с таким же количеством листьев и междоузлий, сжатых до более короткой длины [7, 139].Сходным образом уменьшение длины междоузлий было показано у томатов в ответ на обработку PBZ [140]. Он также заметил, что нанесение PBZ в количестве 400 ppm привело к уменьшению длины междоузлий по сравнению с нанесением PBZ на 200 ppm. PBZ может быть эффективным для получения устойчивых растений и уменьшения высоты растений у некоторых видов без ухудшения качества цветения [141, 142]. Вебстер и Куинлан [143] также сообщили, что PBZ имеет большую эффективность в снижении роста многих видов и сортов фруктов умеренного климата.Аналогичное уменьшение высоты растений было зарегистрировано у картофеля [72], Scaevola [106] и Dianthus caryophyllus [107], Syzygium myrtifolium [123] и Mangifera indica [76] в ответ на обработку PBZ.
Ахмад Назарудин и др. [123] сообщили, что применение PBZ показало задержку роста Syzygium myrtifolium в течение 5-месячного периода после нанесения, в то время как повышенная концентрация PBZ (1,25–3,75 г L –1 ) не приводила к дальнейшему снижению высоты растения.Точно так же Tekalign и Hammes [72] указали, что внесение PBZ как для листвы, так и для увлажнения почвы снижает высоту картофеля, хотя некорневое внесение PBZ более эффективно, чем внесение поливного раствора в почву. Эти авторы далее указали, что увеличение концентрации PBZ (45–90 мг д.в. / растение) привело к уменьшению высоты растения. Кроме того, Yeshitela et al. [76] также описали, что высота манговых деревьев и длина новых побегов сократились из-за внесения PBZ как для листвы, так и для увлажнения почвы.
Обработка PBZ увеличивала толщину коры, размер сосудистых пучков и диаметр сердцевины и приводила к более толстым ножкам [72]. Эта модификация может быть связана с радиальным расширением клеток из-за снижения активности эндогенных ГА в ответ на лечение. Wenzel et al. [144] сообщили, что ГА ограничивает степень радиального расширения органов растений. В стволах двудольных растений изменения формы клеток, по-видимому, вызваны более продольной ориентацией микрофибрилл целлюлозы, откладываемых в стенках клеток, предотвращая расширение параллельно этим микрофибриллам, но позволяя расширение перпендикулярно им [145].Неравномерное распределение и расположение сосудистых элементов в стеблях картофеля привело к неправильной форме стеблей. Различные авторы сообщили о различных результатах у разных видов растений в отношении модификаций анатомии стебля, индуцированных PBZ. PBZ индуцировал как количество, так и длину клеток в стебле сафлора [146]. Берроуз и др. [108] сообщили, что обработка PBZ привела к уменьшению диаметра стебля хризантемы примерно на 50% из-за усиленного развития вторичной ксилемы и заметного уменьшения количества верхушек пучка склеренхимы.В побегах персика PBZ уменьшал долю ксилемы и увеличивал долю флоэмы и коры, а также увеличивал плотность ксилемы [147].
Рост корней
Обработка триазолом индуцировала рост корней огурца, что было связано с повышением уровня эндогенных цитокининов [148]. Обработка PBZ увеличивала длину корней и усиливала боковые корни у растений томатов [94], растений Vigna unguiculata , [149] и Festuca, [150]. PBZ индуцировал рост корней как у кукурузы, так и у пшеницы [151], манго [152], авокадо [153], Abelmoschus esculentus [154] и Ocimum sanctum [101].Свитлик и Миллер [155] сообщили, что длина корня стимулировалась внесением PBZ в низких или умеренных концентрациях. Однако более высокие концентрации могут замедлить рост корней.
PBZ увеличивал диаметр корня за счет увеличения ширины коркового слоя и за счет формирования большего количества вторичных сосудов ксилемы. Эта модификация может быть связана с радикальным ростом клеток из-за снижения активности эндогенных ГА в ответ на лечение [72]. Эти авторы далее указали, что необработанные растения имели более тонкие и длинные корни по сравнению с обработанными растениями.Увеличение диаметра корня коррелирует с более крупными клетками паренхимы коры сои и кукурузы [156]. Увеличение диаметра корня хризантемы произошло за счет увеличения числа рядов и диаметра корковых клеток [108].
PBZ увеличивал диаметр и длину фиброзных корней, усиливал образование боковых корней, уменьшал диаметр сосудов ксилемы; однако ситовые трубки флоэмы показали увеличенный диаметр у обработанных PBZ растений Catharanthus roseus [137].Было обнаружено, что увеличенная длина корня у растений, обработанных PBZ, связана с более крупными клетками паренхимы и способствует расширению клеток в радиальном направлении [7].
Улучшение цветков
PBZ эффективен не только для индукции цветков, но также для индукции цветков в начале и в межсезонье манго [77, 157, 158, 159, 160]. PBZ, ингибитор гиббереллина, снижает уровень вегетативного промотора и тем самым увеличивает соотношение флоригенный промотор / вегетативный промотор, что стимулирует цветение побегов в слабоиндуктивных побегах плодовых культур [76, 161, 162, 163].
Экзогенное применение GA, а также высокие эндогенные уровни гиббереллинов оказались серьезным препятствием на пути дифференциации цветочных почек у ряда фруктов умеренного климата, а также тропических фруктов [164]. PBZ, благодаря своей активности против гиббереллина, может вызывать или усиливать цветение, блокируя превращение каурена в кауреновую кислоту. Последний является предшественником гиббереллинов. PBZ может значительно увеличивать общее фенольное содержание терминальных почек и изменять соотношение флоэмы и ксилемы в стебле [165].Такие изменения могут быть важны для ограничения вегетативного роста и усиления цветения за счет изменения распределения ассимилятов и схемы подачи питательных веществ для нового роста.
Применение PBZ до дифференциации цветочных бутонов или на 3 месяца раньше предполагаемого цветения оказалось эффективным в стимулировании цветения манго без сопутствующего уменьшения длины побегов. Однако более высокая концентрация приводит к уплотнению полога и метелки [166, 167]. Реакция на PBZ варьировалась в зависимости от сорта и нагрузки урожая.Эффективность PBZ в стимулировании цветения Citrus sp. зависит от нагрузки на урожай, так как деревья с высокой плодовой нагрузкой почти не цвели. На деревьях со средней и низкой плодовой нагрузкой PBZ значительно увеличил процент проросших почек и цветочных побегов и уменьшил количество вегетативных побегов [168].
Урожайность плодов и клубней
Урожайность плодов: внекорневая подкормка PBZ (200 частей на миллион) была эффективной для увеличения урожайности и минимизации опадания плодов и растрескивания плодов в ягодах [169]. Эффективность PBZ зависела от стадии развития, поскольку применение PBZ при распускании почек и за 2 недели до цветения винограда значительно увеличивало урожайность [77].
Обработка почвы вокруг ствола дерева (замачивание воротника) было более эффективным, чем внекорневая подкормка, поскольку она обеспечивает надлежащее поглощение и вызывает плодоношение [170]. С другой стороны, Yeshitela [77] сообщил, что применение PBZ как для увлажнения почвы, так и для внекорневой подкормки было эффективным в подавлении вегетативного роста и повышении урожайности манго.
Соуза-Мачадо и др. [171] сообщили, что у растений томатов, обработанных PBZ –, была зарегистрирована значительная ранняя зрелость урожая, но между PBZ и контрольными растениями не было зарегистрировано значительных различий в общей урожайности.Однако Giovinazzo et al. [172] обнаружили значительное увеличение урожая на 13% за счет обработки PBZ вместе с более ранней зрелостью урожая на 6%. Точно так же Берова и Златев [94] сообщили, что применение PBZ повысило как урожайность ранних плодов, так и индекс экономической раннеспелости томатов. Кроме того, Mohamed et al. [173] также сообщили о значительном улучшении урожайности плодов и эффективности использования воды помидорами.
Обычно повышение урожайности манго связано с обработкой PBZ, но Voon et al.[161] подчеркнули важность обеспечения адекватными питательными веществами, орошения и в целом хорошего ухода за деревьями для поддержания этих высоких урожаев. В экспериментах Medonca et al. [174] Кроме того, PBZ увеличил продуктивность «Томми Аткинса». Подобное повышение урожайности манго «Томми Аткинс» (увеличение общего количества плодов и общего веса плодов на дереве) было отмечено в ответ на обработку PBZ [76]. Он также указал, что внесение PBZ в почву увеличивает общее количество плодов на дереве и общий вес плодов на дерево по сравнению с опрыскиванием листьев и повышенным применением PBZ (2.78–8,25 г д.в. / дерево) также привело к более высокому общему количеству плодов на дереве и общему весу плодов на дереве.
Урожайность клубней: В опытах с Balamani и Poovaiah [175] внесение PBZ привело к увеличению урожайности клубней с растения. Однако неясно, были ли указанные приросты урожайности следствием увеличения размера или количества клубней. Напротив, Бандара и Танино [176] сообщили, что PBZ почти удвоил количество клубней на одно растение, не влияя на общий свежий вес клубней.Это несоответствие, вероятно, можно объяснить более холодными условиями выращивания в их эксперименте. Tekalign и Hammes [72] показали, что применение PBZ приводит к снижению количества клубней на растение, что может быть связано с уменьшением количества столонов в результате снижения активности GA, которая может быть связана с инициацией столонов, и сильной отрицательной корреляцией. между свежей массой клубней и числом, означающим, что существенное увеличение размера отдельных клубней было ответственным за прибавку урожая.Это может быть связано с взаимодействием раннего клубнеобразования, повышенным содержанием хлорофилла, повышенной скоростью фотосинтеза и более длительным сохранением фотосинтетически активных листьев в ответ на обработку. Кроме того, эти авторы описали, что применение PBZ увеличивает свежую массу клубней, содержание сухого вещества и удельный вес. Они также указали, что низкая концентрация PBZ увеличивает свежий и сухой вес клубней, в то время как увеличение концентрации PBZ (67,5–90,0 мг д.в. / растение) не увеличивает значения этих параметров.
Аналогичным образом, применение PBZ уменьшило количество клубней картофеля [73, 102], количество мини-клубней [177] и количество клубней маниоки [178]. В отличие от увеличения свежей и сухой массы клубней картофеля [102, 179], мини-клубней картофеля [177] и батата из слоновьих лап [180], свежий вес клубней маниоки был снижен в ответ на обработку PBZ [178].
Качество фруктов и клубней
Качество плодов: Качество плодов манго и лимона (TSS и содержание кислоты) повысилось с применением PBZ [160, 181].Аналогичным образом Виджаялакшми и Шринивасан [111] и Йешитела и др. [76] сообщили, что применение PBZ в манго оказало наибольшее влияние на увеличение общего сахара, снижение содержания сахара, TSS, соотношение сахар: кислота и снижение титруемой кислотности. С другой стороны, PBZ не показал улучшения качества плодов винограда [77], клубники [182] и персика [183].
Внекорневая подкормка 2500–3000 ppm PBZ через три недели после полного цветения повысила уровень TSS и снизила кислотность персика [184] и вишни [185].Yeshitela et al. [76] также сообщили, что применение 8,25 г д.в. / дерево PBZ привело к увеличению TSS. Несмотря на то, что PBZ улучшил качество фруктов, было установлено, что накопление остатков PBZ на поверхности или внутри плода манго (особенно из-за применения более высоких доз) неблагоприятно для здоровья человека [42].
Качество клубней: применение PBZ к растению маниока, значительно увеличившее содержание крахмала, было сообщено Yang и Cao [186] и Medina et al. [178]. Точно так же в картофеле применение PBZ уменьшило разделение ассимилятов на стебель, листья, корень и столон, но увеличило разделение производства сухой массы на клубни [72].Увеличение удельного веса и содержания сухого вещества в клубнях в ответ на PBZ может быть связано со снижением активности GA в ткани клубней, что, в свою очередь, увеличивает силу погружения для привлечения большего количества ассимилятов и усиления синтеза крахмала [72].